Физиология сенсорных систем. Анализаторы. 12. физиология сенсорных систем (анализаторы)
 Скачать 2.54 Mb. Скачать 2.54 Mb.
|

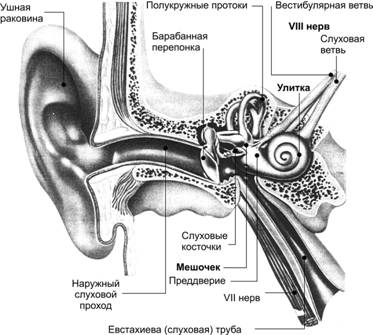 Рис. 12.10. Органы слуха и равновесия. Наружное, среднее и внутреннее ухо, а также отходящие от рецепторных элементов органа слуха (кортиев орган) и равновесия (гребешки и пятна) слуховая и преддверная (вестибулярная) ветви преддверно–улиткового нерва (VIII пара черепных нервов). Механизм передачи и восприятия звука. Звуковые колебания улавливаются ушной раковиной и по наружному слуховому проходу передаются барабанной перепонке, которая начинает колебаться в соответствии с частотой звуковых волн. Колебания барабанной перепонки передаются цепи косточек среднего уха и при их участии мембране овального окна. Колебания мембраны окна преддверия передаются перилимфе и эндолимфе, что вызывает колебания основной мембраны вместе с расположенным на ней кортиевым органом. При этом волосковые клетки своими волосками касаются покровной (текториальной) мембраны, и вследствие механического раздражения в них возникает возбуждение, которое передаётся далее на волокна преддверно-улиткового нерва (рис. 12.11). 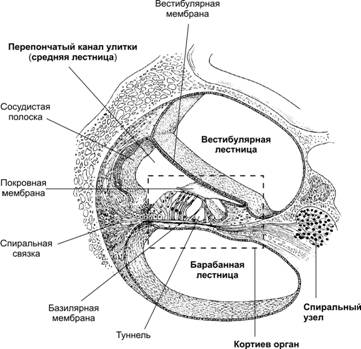 Рис. 12.11. Перепончатый канал и спиральный (кортиев) орган. Канал улитки разделён на барабанную и вестибулярную лестницы и перепончатый канал (средняя лестница), в котором расположен кортиев орган. Перепончатый канал отделён от барабанной лестницы базилярной мембраной. В её составе проходят периферические отростки нейронов спирального ганглия, образующие синаптические контакты с наружными и внутренними волосковыми клетками. Расположение и структура рецепторных клеток кортиевого органа. На основной мембране расположены два вида рецепторных волосковых клеток: внутренние и наружные, отделённые друг от друга кортиевыми дугами. Внутренние волосковые клетки располагаются в один ряд; общее число их по всей длине перепончатого канала достигает 3 500. Наружные волосковые клетки располагаются в 3-4 ряда; их общее число 12 000-20 000. Каждая волосковая клетка имеет удлинённую форму; один её полюс фиксирован на основной мембране, второй находится в полости перепончатого канала улитки. На конце этого полюса есть волоски, или стереоцилии. Их число на каждой внутренней клетке составляет 30-40 и они очень короткие – 4-5 мкм; на каждой наружной клетке число волосков достигает 65-120, они тоньше и длиннее. Волоски рецепторных клеток омываются эндолимфой и контактируют с покровной (текториальной) мембраной, которая по всему ходу перепончатого канала расположена над волосковыми клетками. Механизм слуховой рецепции. При действии звука основная мембрана начинает колебаться, наиболее длинные волоски рецепторных клеток (стереоцилии) касаются покровной мембраны и несколько наклоняются. Отклонение волоска на несколько градусов приводит к натяжению тончайших вертикальных нитей (микрофиламентов), связывающих между собой верхушки соседних волосков данной клетки. Это натяжение чисто механически открывает от 1 до 5 ионных каналов в мембране стереоцилии. Через открытый канал в волосок начинает течь калиевый ионный ток. Сила натяжения нити, необходимая для открытия одного канала, ничтожна, около 2·10-13 ньютон. Ещё более удивительным кажется то, что наиболее слабые из ощущаемых человеком звуков растягивают вертикальные нити, связывающие верхушки соседних стереоцилий, на расстояние, вдвое меньшее, чем диаметр атома водорода. Тот факт, что электрический ответ слухового рецептора достигает максимума уже через 100-500 мкс (микросекунд), означает, что ионные каналы мембраны открываются непосредственно механическим стимулом без участия вторичных внутриклеточных посредников. Это отличает механорецепторы от значительно медленнее работающих фоторецепторов. Деполяризация пресинаптического окончания волосковой клетки приводит к выходу в синаптическую щель нейромедиатора (глутамата или аспартата). Воздействуя на постсинаптическую мембрану афферентного волокна, медиатор вызывает генерацию возбуждения постсинаптического потенциала и далее генерацию распространяющихся в нервных центрах импульсов. Открытие всего нескольких ионных каналов в мембране одной стереоцилии явно мало для возникновения рецепторного потенциала достаточной величины. Важным механизмом усиления сенсорного сигнала на рецепторном уровне слуховой системы является механическое взаимодействие всех стереоцилий (около 100) каждой волосковой клетки. Оказалось, что все стереоцилии одного рецептора связаны между собой в пучок тонкими поперечными нитями. Поэтому, когда сгибается один или несколько более длинных волосков, они тянут за собой все остальные волоски. В результате этого открываются ионные каналы всех волосков, обеспечивая достаточную величину рецепторного потенциала. Бинауральный слух. Человек и животные обладают пространственным слухом, т.е. способностью определять положение источника звука в пространстве. Это свойство основано на наличии двух симметричных половин слухового анализатора (бинауральный слух). Острота бинаурального слуха у человека очень высока: он способен определять расположение источника звука с точностью порядка 1 углового градуса. Физиологической основой этого служит способность нейронных структур слухового анализатора оценивать интерауральные (межушные) различия звуковых стимулов по времени их прихода на каждое ухо и по их интенсивности. Если источник звука находится в стороне от средней линии головы, звуковая волна приходит на одно ухо несколько раньше и большей силы, чем на другое. Оценка удалённости звука от организма связана с ослаблением звука и изменением его тембра. Вестибулярная система Вестибулярная система играет наряду со зрительной и соматосенсорной системами ведущую роль в пространственной ориентировке человека. Она получает, передаёт и анализирует информацию об ускорениях или замедлениях, возникающих в процессе прямолинейного или вращательного движения, а также при изменении положения головы в пространстве. При равномерном движении или в условиях покоя рецепторы вестибулярной сенсорной системы не возбуждаются. Импульсы от вестибулорецепторов вызывают перераспределение тонуса скелетной мускулатуры, что обеспечивает сохранение равновесия тела. Эти влияния осуществляются рефлекторным путём через ряд отделов ЦНС. Периферическим отделом вестибулярного анализатора является вестибулярный аппарат, расположенный в лабиринте пирамиды височной кости. Строение вестибулярного аппарата представлено на схеме 12.1. и рис.12.12. Схема 12.1. Лабиринт (вестибулярный аппарат) Возбуждаются волосковые (рецепторные) клетки Вызывает движение эндолимфы в лабиринте Вызывают смещение отолитов (кристаллики карбоната кальция) Ускоренное движение Повороты и наклоны головы Отолитов аппарат (состоит из двух мешочков - овального и круглого), заполненный желеобразной массой 3 полукружных канала, расположенных взаимно перпендикулярно и заполненных эндолимфой Возбуждение волосковых (рецепторных) клеток происходит вследствие скольжения отолитовой мембраны по волоскам, т.е. их сгибания 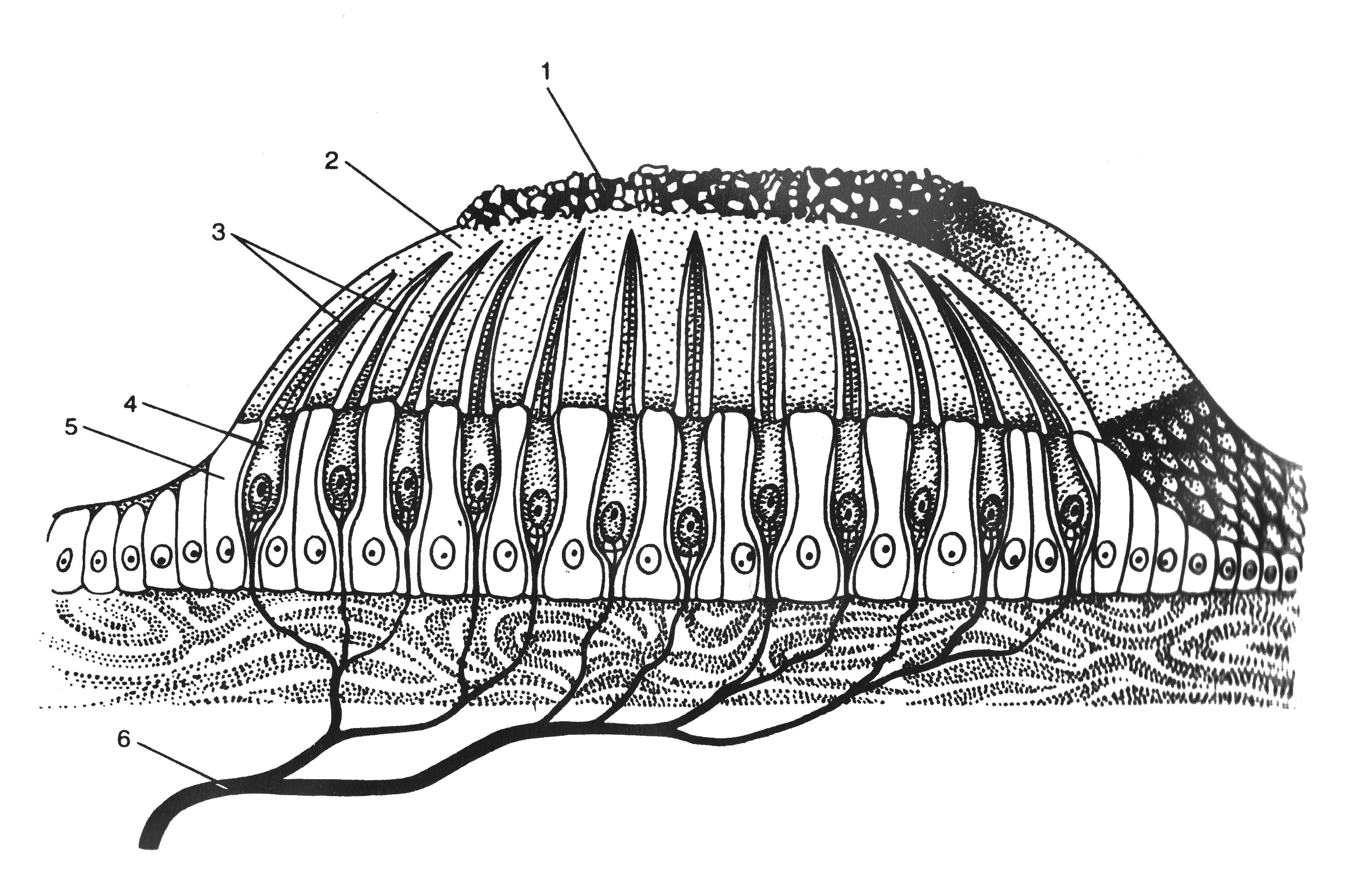 Рис. 12.12. Строение отолитового аппарата. 1 – отолиты; 2 – отолитовая мембрана; 3 – волоски рецепторных клеток; 4 – рецепторные клетки; 5 – опорные клетки; 6 – нервные волокна. При генерации потенциала действия на рецепторных клетках осуществляется его передача через синапс на волокна вестибулярного нерва. Волокна вестибулярного нерва (отростки биполярных нейронов) направляются в продолговатый мозг. Импульсы, проходящие по этим волокнам, поступают на нейроны бульбарного вестибулярного комплекса (ядра Бехтерева, Дейтерса и Швальбе). Отсюда сигналы направляются во многие отделы ЦНС: спинной мозг, мозжечок, глазодвигательные ядра, кору большого мозга, ретикулярную формацию и вегетативные ганглии. Нейроны вестибулярных ядер обладают способностью реагировать на изменение положения конечностей, повороты тела, сигналы от внутренних органов, т.е. осуществлять синтез информации, поступающей от разных источников. При этом они обеспечивают контроль и управление различными двигательными реакциями. Важнейшие из этих реакций - вестибулоспинальные, вестибуловегетативные и вестибулоглазодвигательные. Вестибулоспиналь-ные влияния осуществляются через вестибуло-, ретикуло- и руброспинальные тракты. Таким образом, осуществляется динамическое перераспределение тонуса скелетной мускулатуры и рефлекторные реакции, необходимые для сохранения равновесия. Мозжечок при этом ответствен за фазический характер этих реакций: после его удаления вестибулоспинальные влияния становятся по преимуществу тоническими. Во время произвольных движений вестибулярные влияния на спинной мозг ослабляются. В вестибуловегетативные реакции вовлекаются сердечно-сосудистая система, ЖКТ и другие органы. При сильных длительных нагрузках на вестибулярный аппарат возникает патологический симптомокомплекс, названный болезнью движения, например морская болезнь. Она проявляется изменением сердечного ритма (учащение, а затем замедление), сужением, а затем расширением сосудов, усилением сокращений желудка, головокружением, тошнотой и рвотой. Повышенная склонность к болезни движения может быть уменьшена специальной тренировкой (вращение, качели) и применением ряда лекарственных средств. Вестибулоглазодвигательные рефлексы (глазной нистагм) состоят в медленном движении глаз в противоположную вращению сторону, сменяющимся скачком глаз обратно. Само возникновение и характеристика вращательного глазного нистагма – важные показатели вестибулярной системы, они широко используются в морской, авиационной и космической медицине, а также в эксперименте и клинике. Центральный отдел вестибулярной сенсорной системы расположен в височной доле коры больших полушарий. Вестибулярный анализатор помогает организму ориентироваться в пространстве при активном движении или при пассивном переносе с места на место с завязанными глазами. При этом лабиринтный аппарат с помощью корковых отделов системы анализирует и запоминает направление движения, повороты и пройденное расстояние. Следует подчеркнуть, что в нормальных условиях пространственная ориентировка обеспечивается совместной деятельностью зрительной и вестибулярной сенсорных систем. Обонятельный анализатор Рецепторы обонятельного анализатора расположены в области верхних носовых ходов (рис. 12.13). Обонятельный эпителий находится в стороне от дыхательного пути, он имеет толщину 100-150 мкм и содержит рецепторные клетки диаметром 5-10 мкм, расположенные между опорными клетками. Общее число обонятельных рецепторов человека – около 10 млн (рис. 12.14). 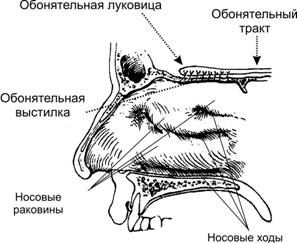 Рис. 12.13. Топография обонятельной выстилки, обонятельной луковицы и обонятельного тракта На поверхности каждой обонятельной клетки имеется сферическое утолщение – обонятельная булава, из которой выступает по 6-12 тончайших волосков длинной до 10 мкм. Обонятельные волоски погружены в жидкую среду, вырабатываемую боуменовыми железами. 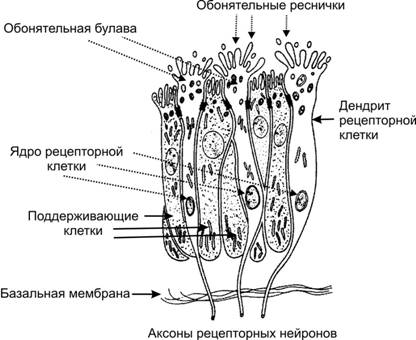 Рис. 12.14. Строение обонятельного эпителия Считается, что наличие волосков в десятки раз увеличивает площадь контакта рецептора с молекулами пахучих веществ. Не исключена и активная, двигательная функция волосков, увеличивающая надёжность захвата молекул пахучего вещества и контакта с ним. Продолжительность жизни обонятельной клетки около 2 мес. Молекулы пахучего вещества вступают в контакт со слизистой оболочкой носовых ходов, взаимодействуют со специализированными белками, встроенными в мембрану рецептора. В результате следующей за этим сложной и пока ещё недостаточно изученной цепи реакций в рецепторе генерируется рецепторный потенциал, а затем и импульсное возбуждение, передающееся по волокнам обонятельного нерва в обонятельную луковицу – первичный нервный центр обонятельного анализатора. Особенность обонятельного анализатора состоит в том, что его афферентные волокна не переключаются в таламусе и не переходят на противоположную сторону большого мозга. Выходящий из луковицы обонятельный тракт состоит из нескольких пучков, которые направляются в разные отделы переднего мозга: переднее обонятельное ядро, обонятельный бугорок, препериформную кору, периамигдалярную кору и часть ядер миндалевидного комплекса. Связь обонятельной луковицы с гиппокампом, периформной корой и другими отделами обонятельного мозга осуществляется через несколько переключений. Многочисленные исследования показали, что наличие значительного числа центров обонятельного мозга не является необходимым для опознания запахов. Поэтому большинство областей проекции обонятельного тракта можно рассматривать как ассоциативные центры, обеспечивающие связь обонятельной системы с другими сенсорными системами и организацию на этой основе ряда сложных форм поведения – пищевой, оборонительной, половой и т.д. Чувствительность обонятельного анализатора человека чрезвычайно велика: один обонятельный рецептор может быть возбуждён одной молекулой пахучего вещества, а возбуждение небольшого числа рецепторов приводит к возникновению ощущения. В то же время изменение интенсивности действия вещества (порог различения) оценивается людьми довольно грубо (наименьшее воспринимаемое различие в силе запаха составляет 30-60% от его исходной концентрации). У собак эти показатели в 3-6 раз меньше. Вкусовой анализатор В процессе эволюции вкус формировался как механизм выбора или отвергания пищи. В естественных условиях вкусовые ощущения комбинируются с обонятельными, тактильными и термическими, также создаваемыми пищей. Важным обстоятельством является то, что предпочтительный выбор пищи отчасти основан на врождённых рефлексах, но в значительной мере на выработанных в онтогенезе условнорефлекторным путём. 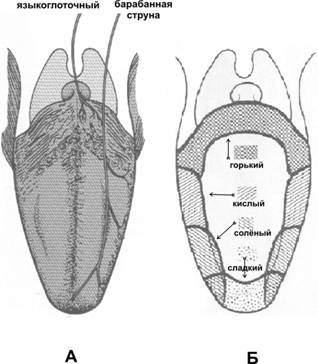 Рис. 12.15. Вкусовые области языка. Вкус сладкого регистрируется преимущественно на кончике языка, солёного — ближе к кончику языка, кислого — на боковых сторонах языка, горького — в задней части языка и в мягком нёбе. Вкус, так же как и обоняние, основан на хеморецепции. Вкусовые рецепторы несут информацию о характере и концентрации веществ, поступающих в рот. Их возбуждение запускает сложную цепь реакций разных отделов мозга, приводящих к работе органов пищеварения или удалению вредных для организма веществ, попавших в рот с пищей. Вкусовые почки – рецепторы вкуса – расположены на языке (рис. 12.15), задней стенке глотки, мягком нёбе, миндалинах и надгортаннике. Больше всего их на кончике языка, его краях и задней части. Каждая из примерно 10 000 вкусовых почек человека состоит из нескольких (2-6) рецепторных клеток и опорных клеток. Вкусовая почка имеет колбовидную форму (рис. 12.16). Она не достигает поверхности слизистой оболочки языка и соединена с полостью рта через вкусовую пору. 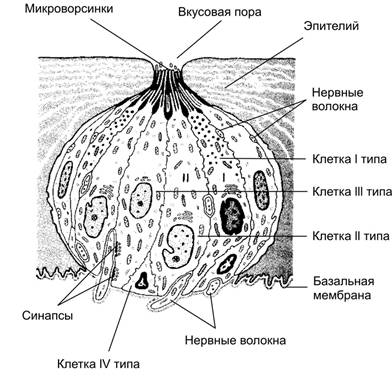 Рис. 12.16. Вкусовая почка состоит из удлинённых светлых клеток различных типов. На верхушечной (апикальной) части большинства клеток расположены микроворсинки — вкусовые волоски, занимающие вкусовой канал, открывающийся на поверхность эпителия вкусовой порой. В микроворсинки «вмонтированы» рецепторные вкусовые белки. В базальной части вкусовой луковицы рецепторные клетки образуют синапсы с терминальными разветвлениями периферических отростков первичных чувствительных нейронов. Каждая вкусовая почка иннервирована примерно 50 нервными волокнами, а каждый чувствительный нейрон получает сигналы примерно от 5 вкусовых луковиц. Вкусовые клетки – наиболее короткоживущие эпителиальные клетки организма; в среднем через каждые 250 часов каждая клетка сменяется молодой. Каждая из рецепторных клеток имеет тончайшие микроворсинки. Считают, что они играют важную роль в возбуждении рецепторной клетки, воспринимая те или иные химические вещества, адсорбированные в канале почки. Этапы первичного преобразования химической энергии вкусовых веществ в энергию нервного возбуждения вкусовых рецепторов ещё не известны. Проводниками всех видов вкусовой чувствительности служат барабанная струна и языкоглоточный нерв, ядра которых в продолговатом мозге содержат первые нейроны вкусового анализатора. Регистрация импульсации в отдельных волокнах данных нейронов показала, что многие из волокон отличаются определённой специфичностью, так как отвечают лишь на соль, кислоту, хинин или сахар. Однако наиболее убедительной считается гипотеза, согласно которой информация о 4 основных вкусовых ощущениях: горьком, сладком, кислом и солёном – кодируется не импульсацией в одиночных волокнах, а разным распределением частоты разрядов в большой группе волокон одновременно, но по-разному возбуждаемых вкусовым веществом. Вкусовые афферентные сигналы поступают в ядро одиночного пучка ствола мозга. От ядра одиночного пучка аксоны вторых нейронов входят в состав медиальной петли до дугообразного ядра таламуса, где расположены третьи нейроны, дающие аксоны до корковых центров вкуса. В настоящее время результаты исследований ещё не позволяют оценить характер преобразований вкусовых афферентных сигналов на всех уровнях вкусового анализатора. У разных людей абсолютные пороги вкусовой чувствительности к разным веществам могут существенно отличаться вплоть до «вкусовой слепоты» к отдельным агентам (например, к креатину). Кроме того, абсолютные пороги вкусовой чувствительности во многом зависят от состояния организма (они изменяются при голодании, беременности и т.д.). При измерении абсолютной вкусовой чувствительности возможны две её оценки: возникновение неопределённого вкусового ощущения (отличающегося от вкуса дистиллированной воды) и возникновение определенного вкусового ощущения. Порог возникновения второго ощущения выше. Пороги различения минимальны в диапазоне средних концентраций веществ, но при переходе к большим концентрациям резко повышаются. Поэтому 20% раствор сахара воспринимается как максимально сладкий, 10% раствор хлорида натрия – как максимально солёный, 0,2% раствор соляной кислоты – максимально кислый, а 0,1% раствор сульфата хинина – максимально горький. Пороговый контраст для разных веществ значительно колеблется. При действии вкусовых веществ наблюдается адаптация (снижение интенсивности вкусового ощущения). Продолжительность адаптации пропорциональна концентрации раствора. Адаптация к сладкому и солёному развивается быстрее, чем к горькому и кислому. Обнаружена и перекрёстная адаптация, т.е. изменение чувствительности к одному веществу при действии другого. Применение нескольких вкусовых раздражителей одновременно или последовательно даёт эффекты вкусового контраста или смещения вкуса. Например, адаптация к горькому повышает чувствительность к кислому и солёному, адаптация к сладкому обостряет восприятие всех других вкусовых ощущений. При смешении нескольких вкусовых веществ может возникнуть новое вкусовое ощущение, отличающееся от вкуса составляющих смесь компонентов. |