2. Строение и функции белков
 Скачать 0.73 Mb. Скачать 0.73 Mb.
|

|
Вторичная структура ДНК молекула ДНК имеет форму спирали, образованную двумя полинуклеотидными цепями, закрученными относительно друг друга и вокруг общей оси. Двойная спираль правозакрученная, полинуклеотидньхе цепи в ней антипараллельны(рис. 4-6), т.е. если одна из них ориентирована в направлении 3'→5', то вторая - в направлении 5'→3'. Третичная структура ДНК (суперспирализация ДНК) Каждая молекула ДНК упакована в отдельную хромосому. В диплоидных клетках человека содержится 46 хромосом. Общая длина ДНК всех хромосом клетки составляет 1,74 м. 39.Моносахариды, строение. Моносахариды - производные многоатомных спиртов, содержащие карбонильную группу. В зависимости от положения в молекуле карбонильной группы моносахариды подразделяют на альдозы и кетозы.Альдозы содержат функциональную альдегидную группу -НС=О, тогда как кетозы содержат кетонную группу >С=О. Название моносахарида зависит от числа составляющих его углеродных атомов, например альдотриозы, кетотриозы, альдогексозы, кетогексозы и т.д.Моносахариды по строению можно отнести к простым углеводам, так как они не гидролизуются при переваривании, в отличие от сложных, которые при гидролизе распадаются с образованием простых углеводов. Строение основных представителей моносахаридов показано на рис. 7-1.В пище человека (фрукты, мёд, соки) содержится небольшое количество моносахаридов, в основном глюкоза и фруктоза. Глюкоза является альдогексозой. Она может существовать в линейной и циклической формах. Циклическая форма глюкозы, предпочтительная в термодинамическом отношении, обусловливает химические свойства глюкозы. Как и все гексозы, глюкоза имеет 4 асимметричных углеродных атома, обусловливающих наличие сте-реоизомеров. Возможно образование 16 стереоизомеров, наиболее важные из которых D- и L-глюкоза. Эти типы изомеров зеркально отображают друг друга (рис. 7-2). Расположение Н- и ОН-групп относительно пятого углеродного атома определяет принадлежность глюкозы к D- или L-ряду. В организме млекопитающих моносахариды находятся в D-конфигурации, так как к этой форме глюкозы специфичны ферменты, катализирующие её превращения. В растворе при образовании циклической формы моносахарида образуются ещё 2 изомера (α- и β-изомеры), называемые аномерами, обозначающие определённую конформа-цию Н- и ОН-групп относительно С, (рис. 7-3). У α-D-глюкозы ОН-группа располагается ниже плоскости кольца, а у β-D-глюкозы, наоборот, над плоскостью кольца. Реакции моносахаридов. Присутствие гидроксильных, альдегидных и кетонных групп позволяет моносахаридам вступать в реакции, характерные для спиртов, альдегидов или кетонов. Эти реакции довольно многочисленны. В данном разделе будут описаны лишь некоторые из них, причём в основном имеющие наибольшее биологическое значение.В этом разделе основные реакции моносахаридов рассмотрены на примере D-глюкозы (рис. 7-5), хотя надо иметь в виду, что в метаболизме углеводов принимают участие и другие моносахариды, а также их производные. Мутаротация, или аномеризация - взаимопревращение аномерных форм моносахаридов, α- и β-формы аномеров находятся в растворе в состоянии равновесия. При достижении этого равновесия происходит мутаротация - размыкание и замыкание пиранового кольца и, соответственно, изменение расположения Н- и ОН-групп при первом углероде моносахарида. Образование гликозидов. Гликозидная связь имеет важное биологическое значение, потому что именно с помощью этой связи осуществляется ковалентное связывание моносахаридов в составе олиго- и полисахаридов. При образовании гликозидной связи аномерная ОН-группа одного моносахарида взаимодействует с ОН-группой другого 40.Дисахариды Дисахариды - это углеводы, молекулы которых состоят из двух остатков моносахаридов, соединенных друг с другом за счет взаимодействия гидроксильных групп (двух полуацетальных или одной полуацетальной и одной спиртовой). Примером наиболее распространенных в природе дисахаридов является сахароза (свекловичный или тростниковый сахар). Молекула сахарозы состоит из остатков глюкозы и фруктозы, соединенных друг с другом за счет взаимодействия полуацетальных гидроксилов. 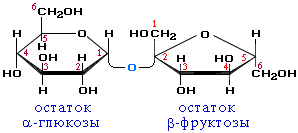 Сахароза, находясь в растворе, не вступает в реакцию "серебряного зеркала", так как не способна образовывать открытую форму. Подобные дисахариды называют невосстанавливающими, т.е. не способными окисляться. Существуют дисахариды, в молекулах которых имеется свободный полуацетальный гидроксил, в водных растворах таких сахаров существуют равновесие между открытой и циклической формами молекул. Такие дисахариды легко окисляются, т.е. являются восстанавливающими, например, мальтоза. 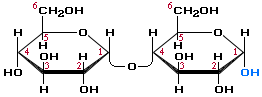 Для дисахаридов характерна реакция гидролиза, в результате которой образуются две молекулы моносахаридов: C12H22O11 + H2O 2C6H12O6 По типу дисахаридов построены молекулы других олигосахаридов и полисахаридов. Строение и свойства важнейших природных полисахаридов крахмала и целлюлозы рассмотрены в части VI "Высокомолекулярные соединения" 41.Полисахариды. Строение крахмала. Полисахариды —класс сложных высокомолекулярных углеводов, молекулы которых состоят из мономеров — моносахаридов. Важнейшие представители несахароподобных полисахаридов – крахмал и целлюлоза (клетчатка). Эти углеводы во многом отличаются от моно- и олигосахаридов. Они не имеют сладкого вкуса, большинство из них не растворимо в воде. По этой причине их называют не сахароподобными . Несахароподобные полисахариды представляют собой высокомолекулярные соединения, которые под каталитическим влиянием кислот или ферментов подвергаются гидролизу с образованием более простых полисахаридов, затем дисахаридов и, в конечном итоге, множества (сотен и тысяч) молекул моносахаридов. Химическое строение полисахаридов. По химической природе полисахариды стоит рассматривать как полигликозиды(полиацетали). Каждое звено моносахарида связано гликозидными связями с предыдущим и последующим звеньями. При этом для связи с последующим звеном предоставляется полуацетальная(гликозидная) гидроксильная группа, а с предыдущим – спиртовая гидроксильная группа. 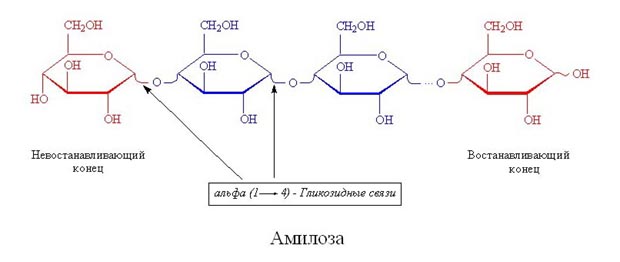 На конце цепи находится остаток восстанавливающегося моносахарида. Но поскольку доля концевого остатка относительно всей макромолекулы весьма невелика, то полисахариды проявляют очень слабые восстановительные свойства. Гликозидная природа полисахаридов обусловливает их гидролиз в кислой и высокую устойчивость в щелочной средах. Полисахариды имеют большую молекулярную массу. Им присущ характерный для высокомолекулярных веществ более высокий уровень структурной организации макромолекул. Наряду с первичной структурой, т.е. определённой последовательностью мономерных остатков, важную роль играет вторичная структура, определяемая пространственным расположением молекулярной цепи. Классификация полисахаридов. Полисахариды можно классифицировать по разным признакам. Полисахаридные цепи могут быть: разветвлёнными или неразветвлёнными (линейными). Также, различают: гомополисахаридами - полисахариды, состоящие из остатков одного моносахарида, гетерополисахариды - полисахариды, состоящие из остатков разных моносахаридов. Наиболее изучены гомополисахариды. Их можно разделить по их происхождению: гомополисахариды растительного происхождения - Крахмалл, - Целюлоза, - Пектиновые вещества и т.д. Гетерополисахариды, к числу которых относятся многие животные и бактериальные полисахариды, изучены меньше, однако они играют важную биологическую роль. Гетерополисахариды в организме связаны с белками и образуют сложные надмолекулярные комплексы. Для полисахаридов используется общее название гликаны. Гликаны могут быть: гексозанами (состоят из гексоз), пентозанами, (состоят из пентоз). В зависимости от природы моносахарида различают: глюканы (в основе – моносахарид глюкоза), маннаны (в основе – моносахарид манноза), галактаны (в основе – моносахарид галактоза) и т.п. Крахмал Крахмал (С6Н10О5)n – белый (под микроскопом зернисый) порошок, нерастворимый в холодной воде. В горячей воде крахмал набухает, образуя коллоидный раствор (крахмальный клейстер). С раствором йода даёт синее окрашивание (характерная реакция). Крахмал образуется в результате фотосинтеза, в листьях растений, и запасается в клубнях, корнях, зёрнах. Химическое строение крахмала Крахмал представляет собой смесь двух полисахаридов, построенных из глюкозы (D-глюкопиранозы): амилозы (10-20%) и амилопектина (80-90%). Дисахаридным фрагментом амилозы является мальтоза. В амилозе D-глюкопиранозные остатки связаны альфа(1-4) гликозидными связями. 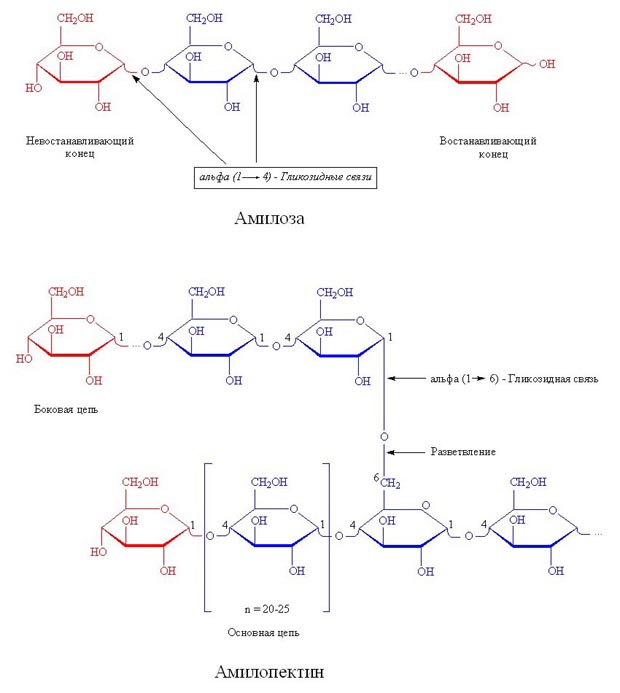 42. Переваривание углеводов. Эпителиальные клетки кишечника способны всасывать только моносахариды. Поэтому процесс переваривания заключается в ферментативном гидролизе гликозидных связей в углеводах, имеющих олиго- или полисахаридное строение 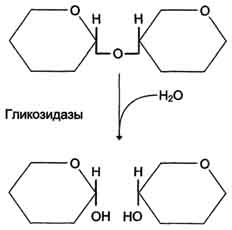 А. Переваривание углеводов в ротовой полости В ротовой полости пища измельчается при пережёвывании, смачиваясь при этом слюной. Слюна на 99% состоит из воды и обычно имеет рН 6,8. В слюне присутствует гидролитический фермент α-амилаза (α-1,4-гликозидаза), расщепляющая в крахмале α-1,4-гликозидные связи. В ротовой полости не может происходить полное расщепление крахмала, так как действие фермента на крахмал кратковременно. Кроме того, амилаза слюны не расщепляет α- 1,6-гликозидные связи (связи в местах разветвлений), поэтому крахмал переваривается лишь частично с образованием крупных фрагментов - декстринов и небольшого количества мальтозы. Следует отметить, что амилаза слюны не гидролизует гликозидные связи в дисахаридах. Действие амилазы слюны прекращается в резко кислой среде содержимого желудка (рН 1,5-2,5). Однако внутри пищевого комка активность амилазы может некоторое время сохраняться, пока рН не изменится в кислую сторону. Желудочный сок не содержит ферментов, расщепляющих углеводы. В желудочном содержимом возможен лишь незначительный кислотный гидролиз гликозидных связей. Б. Переваривание углеводов в кишечнике Последующие этапы переваривания нерасщеплённого или частично расщеплённого крахмала, а также других углеводов пищи происходит в тонком кишечнике в разных его отделах под действием гадролитических ферментов - гликозидаз. Панкреатическая α-амилаза В двенадцатиперстной кишке рН среды желудочного содержимого нейтрализуется, так как секрет поджелудочной железы имеет рН 7,5-8,0 и содержит бикарбонаты (НСО3-). С секретом поджелудочной железы в кишечник поступает панкреатическая α-амилаза. Этот фермент гидролизует α-1,4-гликозидные связи в крахмале и декстринах. Продукты переваривания крахмала на этом этапе - дисахарид мальтоза, содержащая 2 остатка глюкозы, связанные α-1,4-связью. Из тех остатков глюкозы, которые в молекуле крахмала находятся в местах разветвления и соединены α-1,6-гликозидной связью, образуется дисахарид изомальтоза. Кроме того, образуются олигосахариды, содержащие 3-8 остатков глюкозы, связанные α-1,4- и α-1,6-связями (рис. 7-11). α-Амилаза поджелудочной железы, так же, как α-амилаза слюны, действует как эндогликозидаза. Панкреатическая α-амилаза не расщепляет α-1,6-гликозидные связи в крахмале. Этот фермент также не гидролизует (3-1,4-гликозидные связи, которыми соединены остатки глюкозы в молекуле целлюлозы. Целлюлоза, таким образом, проходит через кишечник неизменённой. Тем не менее непереваренная целлюлоза выполняет важную функцию балластного вещества, придавая пище дополнительный объём и положительно влияя на процесс переваривания. Кроме того, в толстом кишечнике целлюлоза может подвергаться действию бактериальных ферментов и частично расщепляться с образованием спиртов, органических кислот и СО2. Продукты бактериального расщепления целлюлозы важны как стимуляторы перистальтики кишечника. Особенность переваривания углеводов в тонком кишечнике заключается в том, что активность специфических олиго- и дисахаридаз в просвете кишечника низкая. Но ферменты активно действуют на поверхности эпителиальных клеток кишечника. Ферменты, расщепляющие гликозидные связи в дисахаридах (дисахаридазы), образуют ферментативные комплексы, локализованные на наружной поверхности цитоплазматической мембраны энтероцитов. 43. Этапы гликолиза. Первый этап гликолиза– подготовительный, здесь происходит затрата энергии АТФ, активация глюкозы и образование из нее триозофосфатов.Первая реакциягликолиза сводится к превращению глюкозы в реакционно-способное соединение за счет фосфорилирования 6-го, не включенного в кольцо, атома углерода. Эта реакция является первой в любом превращении глюкозы, катализируется гексокиназой. Вторая реакциянеобходима для выведения еще одного атома углерода из кольца для его последующего фосфорилирования (фермент изомераза). В результате образуется фруктозо-6-фосфат. Третья реакция– фермент фосфофруктокиназа фосфорилирует фруктозо-6-фосфат с образованием почти симметричной молекулы фруктозо-1,6-дифосфата. Эта реакция является главной в регуляции скорости гликолиза. Вчетвертой реакциифруктозо-1,6-дифосфат разрезается пополам фруктозо-1,6-дифосфат-альдолазой с образованием двух фосфорилированных триоз-изомеров – альдозы глицеральдегида (ГАФ) и кетозы диоксиацетона (ДАФ). Пятая реакция– переход глицеральдегидфосфата и диоксиацетонфосфата друг в друга при участии триозофосфатизомеразы. Равновесие реакции сдвинуто в пользу диоксиацетонфосфата, его доля составляет 97%, доля глицеральдегидфосфата – 3%. Эта реакция, при всей ее простоте, определяет дальнейшую судьбу глюкозы:при нехватке энергии в клетке и активации окисления глюкозы диоксиацетонфосфат превращается в глицеральдегидфосфат, который далее окисляется на втором этапе гликолиза, при достаточном количестве АТФ, наоборот, глицеральдегидфосфат изомеризуется в диоксиацетонфосфат, и последний отправляется на синтез жиров. Второй этап гликолиза– это освобождение энергии, содержащейся в глицеральдегидфосфате, и запасание ее в форме АТФ. Шестая реакция гликолиза (фермент глицеральдегидфосфат-дегидрогеназа) – окисление глицеральдегидфосфата и присоединение к нему фосфорной кислоты приводит к образованию макроэргического соединения 1,3-дифосфоглицериновой кислоты и НАДН. В седьмой реакции(фермент фосфоглицераткиназа) энергия фосфоэфирной связи, заключенная в 1,3-дифосфоглицерате тратится на образование АТФ. Реакция получила дополнительное название – реакция субстратного фосфорилирования, что уточняет источник энергии для получения макроэргической связи в АТФ (от субстрата реакции) в отличие от окислительного фосфорилирования (от электрохимического градиента ионов водорода на мембране митохондрий). Восьмая реакция– синтезированный в предыдущей реакции 3-фосфоглицерат под влиянием фосфоглицератмутазы изомеризуется в 2-фосфоглицерат. Девятая реакция– фермент енолаза отрывает молекулу воды от 2-фосфоглицериновой кислоты и приводит к образованию макроэргической фосфоэфирной связи в составе фосфоенолпирувата. Десятая реакциягликолиза – еще одна реакция субстратного фосфорилирования – заключается в переносе пируваткиназой макроэргического фосфата с фосфоенолпирувата на АДФ и образовании пировиноградной кислоты. Реакция одиннадцатая– образование молочной кислоты из пирувата под действием лактатдегидрогеназы. Важно то, что эта реакция осуществляется только в анаэробных условиях. Эта реакция необходима клетке, так как НАДН, образующийся в 6-й реакции, в отсутствие кислорода не может окисляться в митохондриях. 45. Гликоген. Биологическая роль. Синтез гликогена. Биологическая роль гликогена Глюкоза является для организма главным энергетическим материалом, поддерживающим его основные функции. Печень складирует глюкозу в форме гликогена не столько для своих нужд, сколько для того, чтобы обеспечивать приток глюкозы к другим тканям – главным образом, эритроцитам и мозгу. Как было сказано выше, клетки мышц, подобно клеткам печени, тоже способны обращать глюкозу в гликоген. Однако гликоген, содержащийся в мышцах, тратится только на мышечную работу. Иными словами, гликоген в мышцах остаётся источником глюкозы лишь для самой клетки, в то время как гликоген, запасённый в печени, после переработки в глюкозу тратится на питание всего организма, а главное - на поддержание в крови нужной концентрации глюкозы. Синтез гликогена (гликогенез) Прежде всего глюкоза подвергается фосфорилированию при участии ферментагексокиназы, а в печени – и глюкокиназы. Далее глюкозо-6-фосфат под влиянием фермента фосфоглюкомутазыпереходит в глюкозо-1-фос-фат: 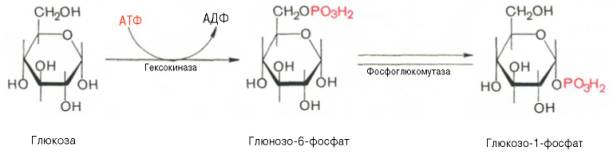 Образовавшийся глюкозо-1-фосфат уже непосредственно вовлекается в синтез гликогена. На первой стадии синтеза глюкозо-1-фосфат вступает во взаимодействие с УТФ (уридинтрифосфат), образуя уридиндифосфатглюкозу (УДФ-глюкоза) и пирофосфат. Данная реакция катализируется ферментом глюкозо-1-фосфат-уридилилтрансферазой (УДФГ-пирофосфорилаза): |