Общая экология (лекции 3-5). 9. структура и динамика популяций понятие о популяции
 Скачать 2.71 Mb. Скачать 2.71 Mb.
|

12.12. Развитие биосферы вноосферу — сферу разумаС появлением человеческого общества, под влиянием которого в современных условиях происходит дальнейшая эволюция биосферы, приводит к изменению качественного состава самой биосферы, к ее переходу в ноосферу. Под ноосферой понимают сферу взаимодействия природы и общества, в котором разумная деятельность людей становится главным, определяющим фактором развития. Название «ноосфера» происходит от греч. «ноос» — разум и таким образом обозначает сферу разума. Понятие ноосферы ввел в 1927 г. французский ученый-математик Э. Леруа, подразумевая под ним современную геологическую стадию развития биосферы. Э. Леруа отмечал, что пришел к такому представлению вместе со своим другом — геологом и палеонтологом П. Тейером де Шарденом, который в дальнейшем разработал собственное представление о ноосфере. В книге «Феномен человека» автор определил ноосферу как «новый покров», «мыслящий пласт, который, зародившись в конце третичного периода, разворачивается над миром растений и животных — вне биосферы и над ней». Научное и практическое значение деятельности В. И. Вернадского как основателя учения о биосфере состоит в том, что он впервые во всеоружии знаний своего времени глубоко обосновал единство человека и биосферы. Сама живая материя как носитель разума, отмечал В. И. Вернадский, составляет небольшую часть биосферы по массе. Возникновение человеческого общества явилось результатом длительного развития живого вещества в пределах биосферы. Появление человека на Земле предопределило неизбежность возникновения нового состояния биосферы — переход ее в ноосферу, оболочку разума, охваченную целенаправленной деятельностью самого человека. При этом периоду сознательной деятельности человека предшествовал длительный период его дикого, полудикого и в целом стихийного существования. В пределах биосферы возникла первоначально сфера первобытной деятельности человеческого общества, которую нередко называют антропос-ферой. Начало ей положило расселение человека по всей поверхности суши в результате использования огня. Человек, овладев огнем, стал относительно независимым от климата и заселил все континенты, кроме Антарктиды. По свидетельству уникальных палеонтологических находок, человек, зародившись в дебрях Центральной Африки, освоил Европу, Азию, Австралию, а при дальнейшем совершенствовании своего организма достиг просторов Северной и Южной Америки. В ходе развития производительных сил антропосфера, охватывающая стихийную деятельность человеческого общества, объективно должна перейти в ноосферу — сферу сознательной деятельности. В современную эпоху становление ноосферы теснейшим образом связано с овладением различными формами движения материи — первоначально механической, потом тепловой, химической, атомно-ядерной. На очереди овладение биологическими формами движения — создание живых форм с помощью методов и средств биотехнологии и генной инженерии. С этим связано и возникновение новых по качеству круговоротов веществ в биосфере. В. И. Вернадский, оценивая роль человеческого разума и научной мысли, делает следующие выводы. 1. Ход научного творчества является той силой, которой человек меняет биосферу, в которой он живет. 2. Это проявление изменения биосферы есть неизбежное явление, сопутствующее росту научной мысли. 3. Это изменение биосферы происходит независимо от че-ловеской воли, стихийно, как природный естественный процесс. 4. А так как среда жизни есть организованная оболочка планеты — биосфера, то вхождение в нее в ходе ее геологически длительного существования нового фактора ее изменения — научной работы человечества — есть природный процесс перехода биосферы в новую фазу, в новое состояние — ноосферу. 5. В переживаемый нами исторический момент мы видим это более ясно, чем могли видеть раньше. Здесь вскрываются перед нами «законы природы». Новые науки — геохимия и биогеохимия —дают возможность впервые выразить некоторые важные черты процесса математически. Выводы о том, что биосфера неизбежно превратится в ноосферу, т. е. сферу, где разум человека будет играть доминирующую роль в развитии системы «человек — природа», получили название закона ноосферы В. И. Вернадского. В дальнейшем эволюции биосферы и переходу ее в ноосферу были посвящены научные работы многих зарубежных и отечественных исследователей. Так, М. М. Камшилов (1974), рассматривая эволюцию биосферы, отмечал (рис. 12.51): 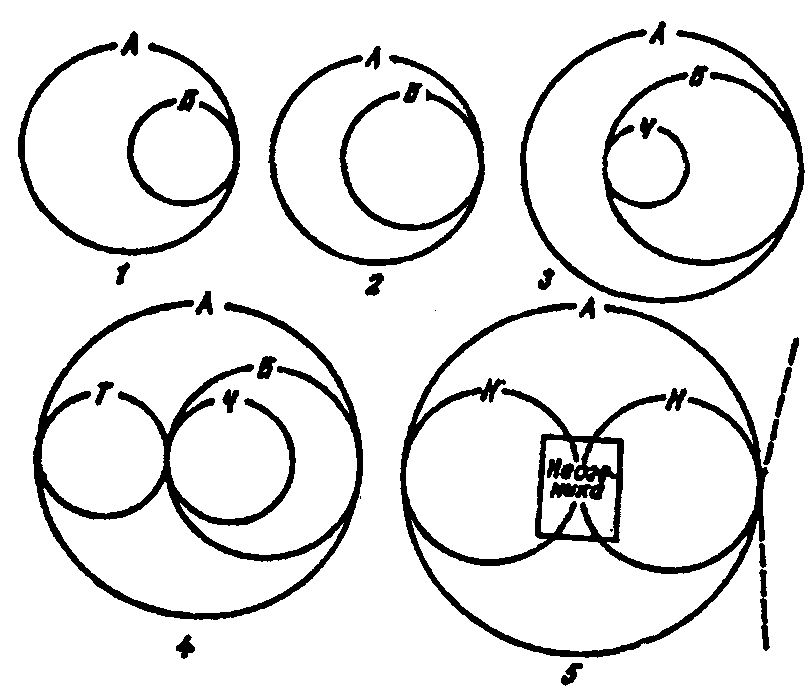 Рис. 12.51. Стадии эволюции биосферы, представленные в виде последовательно вовлекаемых в круговорот циклов (по М. М. Камшилову, 1974) 1 — в большом абиотическом круговороте веществ (А) возникла биосфера (Б); 2 — по мере развития жизни она расширяется; 3 — в ней появляется человеческое общество (Ч); 4 — человеческое общество начинает поглощать вещество и энергию не только через биосферу, но и непосредственно из абиотической среды (Т); 5 — биосфера, превратившаяся в ноосферу (Н), стала развиваться под контролем разумной деятельности человека (ноогенез); управление взаимными отношениями человеческого общества и природы осуществляется с помощью ноогенетики, жизнь, развиваясь по пути ноогенеза, все полнее осваивает вещество, энергию и потенциал информации неживой приро- | ды, распространяясь за пределы Земли (пунктирные линии). В XX в.накопился огромный фактический материал по биосфере, по производственной деятельности человеческого общества. Рождающаяся ноосфера в своих главных проявлениях характеризуется следующими признаками (рис. 12.52). 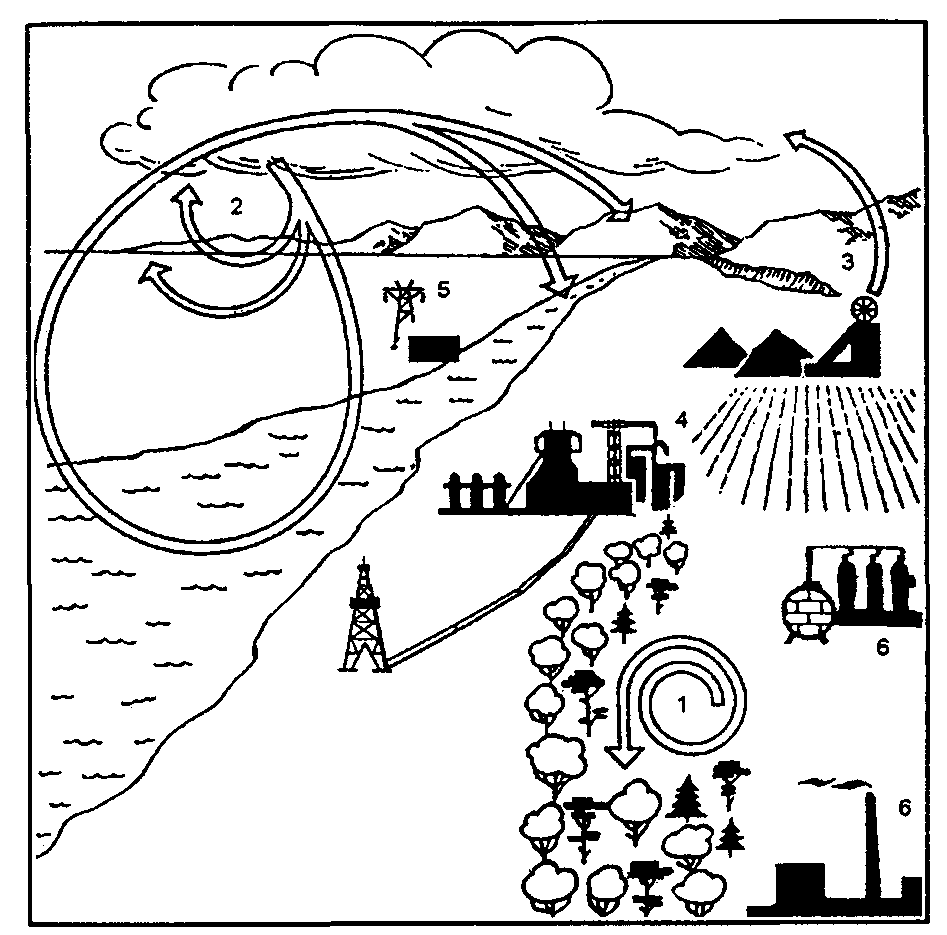 Рис. 12.52. Геохимия ноосферы (по Д. И. Перельману, 1973) Процессы, унаследованные от биосферы, но существенно измененные в ноосфере: 1 — биологический круговорот атомов; 2 — круговорот воды, водная и атмосферная миграция элементов; 3 — рассеяние элементов — отработка месторождений и т. д. Процессы, чуждые биосфере: 4 — получение металлов и других элементов, неустойчивых в термодинамическом поле биосферы; 5 — производство энергии на атомных электростанциях; б — синтез органических веществ, не существовавших в биосфере (полимеры и др.) 1. Возрастающим количеством механически извлекаемого материала литосферы — ростом разработки месторождений полезных ископаемых. В 90-х гг. оно превышало 100 млрд т в год, что в 4 раза больше массы материала, выносимого речным стоком в океан в процессе денудиции суши. 2. Массовым потреблением продуктов фотосинтеза прошлых геологических эпох, главным образом в энергетических целях. Химическое равновесие в биосфере в связи с этим смещается в сторону, противоположную глобальному процессу фотосинтеза, что неизбежно приводит к росту содержания углекислого газа в биосфере и уменьшению содержания свободного кислорода. 3. Процессы в ноосфере приводят к рассеиванию энергии Земли, а не к ее накоплению, что являлось характерным для биосферы до появления человека. Возникает важная энергетическая проблема. 4. В ноосфере создаются в массовом количестве вещества, которые ранее в биосфере отсутствовали. Происходит металлизация биосферы. 5. Характерно для ноосферы появление новых трансурановых химических элементов в связи с развитием ядерной технологии и ядерной энергетики. Овладение ядерной энергией происходит за счет деления тяжелых ядер. Предвидится в недалеком будущем получение термоядерной энергии за счет синтеза легких ядер, что позволит полностью отказаться от горючих полезных ископаемых в качестве источника энергии. 6. Ноосфера выходит за пределы биосферы в связи с огромным прогрессом научно-технической революции. Возникла космонавтика, которая обеспечивает выход человека за пределы планеты Земля. Происходит освоение космического, околокосмического пространства с непредвиденными возможностями. Создается принципиальная возможность создания искусственных биосфер на других планетах. 7. С образованием ноосферы планета Земля переходит в новое качественное состояние. Если биосфера — это сфера Земли, то ноосфера — это сфера Солнечной системы. Ноосфера в будущем станет областью Солнечной системы в познавательных и производственных целях человеческого общества. Таким образом, хаотическое саморазвитие, основанное на процессах естественной саморегуляции, будет заменено разумной стратегией, базирующейся на прогнозно-плановых началах, регулировании процессов естественного развития. Это управление, несомненно, должно быть лишь «мягким» и следовать только законам природы и развития общества. В основе формирования ноосферы могут быть лишь благо и заинтересованное понимание, а не насилие и волюнтаризм. Человечеству придется решать массу тяжелых для нового времени проблем, но это будут иные, чем сегодняшние, проблемы. Целый ряд ученых (В. Л. Барсуков, А. Л. Яншин, 1988; В. А. Кутырев, 1990 и др.) считают закон ноосферы В. И. Вернадского религиозной или социальной утопией. По их мнению, это мечта и вера, конечно, светлая, но весьма далекая от реальности и недостаточная ни как научный прогноз, ни как определение закона ноосферы. Хотя сам закон они считают справедливым. По их мнению, он точен в том смысле, что если человечество не начнет разумно регулировать свою численность и давление на природу в соответствии с ее законами, то в измененном виде биосфера может сохраниться, а цивилизация, не исключено, и вид «человек разумный» погибнут. Развитие по-стантропогенной природы весьма трудно прогнозировать. Одно несомненно, считает Н. Ф. Реймерс (1994), управлять люди будут не природой, а прежде всего собой. И в этом смысл закона ноосферы В. И. Вернадского. Лекция № 8 «Популяционная экология». Органический мир имеет три системных уровня. Отдельный организм – это морфологически структурированная система. Популяций – это информационно-структурированная система, а биогеоценоз, на основе которого, прежде всего, решаются вопросы поддержания биогенного круговорота, – энергетически-структурированная система. Внутривидовая структура. Отдельные особи существуют только как части целого вида. Они объединены в естественные группировки. Само существование вида обеспечивается внутривидовой структурой, основанной на связях между его особями, в том числе на связях в группировках. Характер этих связей сложился в ходе эволюции и прошел по выражению Н.П. Наумова (1965) «горнило естественного отбора». Естественные группировки заселяют определенную территорию ареала и входят в живущие здесь сообщества. Перечисленные параметры каждого вида, как и его внутривидовые связи, возникли в процессе эволюции. Исчезновение одного из видов или появление нового вида меняет круговорот вещества и энергии в местах его обитания. Представление о виде со времени его научного определения Д. Рэем и К. Линнеем существенно изменилось. Биологи 17-18 веков представляли вид как совокупность независимых особей, лишь временно объединяемых в пары, семьи, колонии или стаи. Считалось, что все особи одного вида однотипны по всему ареалу. Но уже в 60-е годы 18 века П.С. Паллас, в 1775 году А.А. Каверзнев и в 1825 году Леопольд фон Бух говорили о возможности образования нового вида путем удаления географической расы (подвида) от видового типа. С появление теории Ч. Дарвина (1859) существование и важность групповой и индивидуальной изменчивости стала очевидной, впоследствии подкрепленная теорией наследственности. Накопление данных привело к замене представления о монотипичном (однородном) виде на представления о виде как системе связанных друг с другом групп особей или популяций. Популяция – это чрезвычайно важная ячейка экосистемы. Именно на ее уровне решаются все вопросы жизнедеятельности вида, именно она является элементарной единицей микроэволюции (эволюции видов), способной реагировать на изменение среды перестройкой своего генофонда. Термин "популяция" был введен В. Иогансеном в 1903 г. Было показано, что популяции отличаются морфо-физиологическими, экологическим и генетическими признаками и занимают либо различные географические районы (подвиды), либо определенные экологические ниши (экотипы, биологические расы – Семенов-Тян-Шанский, 1910; Вавилов, 1931; Гёксли, 1940; Майр, 1947). Низшей систематической единицей большинство биологов полагают подвид, 75% особей которого, заселяя определенную часть ареала, имеют несомненные морфологические отличия («правило 75%»). Каждый подвид в свою очередь распадается на ряд еще более мелких иерархических соподчиненных групп особей или популяций. Среди них различаются территориальные популяции и биологические группировки (расы, экотипы, сезонные фазы и др.). Некоторые из них наследственно обусловлены, некоторые обратимы в первом же поколении. Термин популяция до сих пор вызывает дискуссии и понимается по-разному. Большинство ученых (Арнольди, 1957; Н. Наумов, 1936, 1948, 1956; Беклемишев, 1959, 1960) считают, что популяция – группа особей, которая населяет определенную территорию, входит в состав биоценоза и представляет собой форму существования вида в конкретных ландшафтно-географических условиях. М.С. Гиляров (1954) уточняет, что границы популяции являются границами того биоценоза, в который данный вид входит. Именно этот взгляд на популяцию мы и будем рассматривать. Но следует также добавить, что многие генетики часто под популяцией подразумевают совокупность равно наследственных особей. На самом деле вид, как уже говорилось, очень сложная система различных морфо-физиологических, генетических и экологических группировок территориального, экологического и иерархического характера и внутри себя эти группировки также разнородны. Подвиды занимают географически однородную часть ареала. Они распадаются на географические популяции, заселяющие определенные ландшафтно-географический район со своими ландшафтно-климатическими особенностями. Географические популяции распадаются на экологические популяции, поселения животных в определенных биотопах. Экологическая популяция, если ее территория неоднородна, распадается на мелкие группы, заселяющие мозаичный ландшафт – элементарные популяции, мерусы, демы, парцеллы, внутрипопуляционные группировки. Каждая популяция состоит из нескольких генераций, часто резко отличающихся друг от друга морфо-физиологически и экологически. Далее в состав популяции входят половые группы (самцы, самки), а у общественных насекомых и другие формы с разными функциями. Наконец, благодаря расхождению в разные экологические ниши образуются биологические расы (пищевые расы у насекомых, расы по хозяину у паразитов, яровые и озимые расы рыб). Все эти формы усложняют структуру вида, увеличивают биологическое разнообразие, обогащают формы взаимодействия со средой. Кроме того, внутри вида есть группировки с разными формами использования территории (семьи, стада, колонии). Эти группировки определяют структуру популяций и образ жизни: одиночный, стадный, колониальный. Все в целом создает устойчивость вида в среде обитания. Подвиды. Наиболее крупные территориальные группировки, занимающие географически однородную часть ареала. Они являются низшей систематической единицей. 75% особей подвида, заселяя определенную часть ареала, имеют несомненные морфологические отличия («правило 75%»). В гористых условиях и на островах подвиды обычно занимают меньше площади, чем на равнинах континента. Так зимородок Tanysiptera galatea с Новой Гвинеи и соседних островов занимает в целом маленькую территорию, но образует 8 подвидов. Размеры площади занятой подвидами определяются и подвижностью животных. Так, на небольшой площади всего 6 км2 обнаружено целых 15 подвидов улитки Achatinella mustelina (Гавайские острова). С другой стороны наибольшие размеры территорий подвидов у океанических рыб и континентальных птиц из-за их высокой подвижности. Еще один интересный пример того же плана: у обыкновенной белки площади подвидов в Европейской части России и на юге Сибири, где ландшафты пестры и белки не совершают значительных миграций, очень невелики. Напротив, в монотонной зоне тайги белкам свойственны значительные миграции, а площади их подвидов очень велики. Величина занятой подвидом площади может зависеть и от особенностей внутривидовой организации. Население гусей-гуменников в Евразии распадается на 3 подвида, тогда как серая утка, утка-широконоска и многие другие птицы не только в Евразии, но и в Северной Америке не образуют географических подвидов. Дело в том, что семьи гусей, в отличие от уток, не распадаются после подрастания молодняка и сохраняются до возвращения на родину. Вместе с колониальными гнездовьями это способствует возникновению местных рас даже при отсутствии преград или переходных популяций. Таким образом, появление подвидов зависит не только от мест обитания, но и от экологических особенностей животного. Это подтверждает и клинальная географическая изменчивость, когда тот или иной признак меняется в пространстве постепенно, что не дает возможности выделить подвиды и провести между ними четкие границы. Считается, что клинальная изменчивость отражает градиент изменения среды. Но в ряде случаев она не соответствует изменениям условий существования (неадаптивная клина), и может возникнуть, когда нет постепенно изменяющихся условий. Вероятно, неадаптивная клина связана с подвижностью животных и обменом особями между популяциями. В основе клинальной изменчивости могут лежать и наследственные факторы. Географические популяции. Подвиды часто распадаются на географические популяции, заселяющие определенные ландшафтно-географический район со своими однородными ландшафтно-климатическими особенностями. Их территориальные размеры зависят от особенностей вида и заселенной ими территории. Внутри географической популяции реально осуществляется перекрестное размножение. В результате последнего входящие в ее состав особи обладают общим морфо-физиологическим типом, постоянно или временно (чаще) отличающимся от соседних географических популяций. Кроме того, они обладают единым ритмом жизненных явлений (сроками и темпами размножения, спячки, сезонных миграций и др.), общим ритмом динамики численности, стойкостью против неблагоприятных условий, плодовитостью, оседлостью, перелетностью и т.д. Среди особенностей географических популяций большое значение имеют различия в питании. У животных севера преобладают более калорийные корма, у хищников увеличивается плотоядность. Например, на Кавказе растительные корма встречены в 75% желудков лесной куницы, млекопитающие – в 29%, а птицы – в 70%. В Печеро-Илычском заповеднике частота встреч млекопитающих составляла до 79%, птиц – до 35%, а растительный корм обнаружен всего в 13% желудков. Различны в этих популяциях и способы охоты: кавказские куницы охотятся на земле, а печеро-илычские как на земле, так и на деревьях. Различия в питании могут зависеть не только от климатических условий, но и от обилия и доступности разных видов корма. Воробьиные сычи под Москвой зимой питаются преимущественно млекопитающими, грызунами и насекомоядными, а в Центральной Европе – воробьиными птицами. У северных популяций часто возрастает нетребовательность к пище, эврифагия, и значительная сезонная и годовая изменчивость питания как приспособление к бедной кормовой базе. Кроме питания, у отдельных популяций различаются уровень газообмена и химической терморегуляции, водный обмен и другие эколого-физиологические особенности. Так, у озерных лягушек из дельты Волги, где влажность воздуха выше, проницаемость кожи для воды оказалась 20,2 мм3, а у животных из-под Астрахани, где воздух суше, – 16, 2 мм3. Что касается терморегуляции, то для развития одной генерации сумма эффективных температур меньше на севере. У северных особей и интенсивность кровотока оказалась выше. Географические популяции отличаются плодовитостью и смертностью. Из-за этого их динамика численности не одинакова. Огромное значение в характеристике географических популяций играют особенности размножения. Так, две популяции скандинавской сельди имеют разные нерестилища, отличаются местами зимовок и гнездовой разные популяции скворцов и чаек. У мигрирующих животных большое значение в изоляции популяций принадлежит гнездовому консерватизму. В его основе лежит безусловный рефлекс (инстинкт) и индивидуальный опыт (условный рефлекс). Гнездовой консерватизм описан у огромного числа видов птиц и рыб. В малоблагоприятных частях ареала возникают временные популяции, которые не в состоянии поддержать свое существование размножением, но пополняются пришельцами. Существование одних популяций может зависеть от притока из других популяций. Активная и пассивная эмиграция из оптимальных частей ареала играет важную роль в жизни многих популяций. Пример – популяций камбалы (Platessa platessa) в Балтийском море. Таким образом, существование одних географических популяций может в некоторой степени зависеть от других, пополняясь за счет них постоянно или периодически (чаще) на подъеме численности. Географические популяции разобщены, и их изоляция сопровождается накоплением наследственных различий. Они обладают полной способностью к самовоспроизведению и подобно подвидам могут считаться независимыми популяциями (Беклемишев, 1960). Экологические популяции. Географические популяции распадаются на более мелкие группировки – экологические популяции, поселения животных в определенных биотопах. Они менее постоянны по своему составу и слабо изолированы друг от друга. Экологические популяции входят в состав местных сообществ и образуют разные поселения. Например, у обыкновенной полевки в Московской области есть кустарниково-лесные поселения, размножающиеся с весны и до конца лета и защищенные своим биотопом от высокой смертности (много убежищ, высокий травостой). Численность их стабильна. Напротив, другой вид поселений в сельскохозяйственных угодьях размножается практически круглый год, но нестабильность биотопа приводит к гибели многих животных и к миграциям. Колебания численности этих поселений достигают трехкратных размеров. Данные по большому числу видов показывают, что поселения животных (экологические популяции) можно разделить на постоянные и временные. Первые существуют в благоприятных местах обитания, вторые – в местах с колебаниями условий. Экологические популяции хорошо выделяются у млекопитающих, птиц, пресмыкающихся, земноводных и рыб. У насекомых они выражены менее четко. У прикрепленных и малоподвижных животных они легко выделяются даже на небольшой территории. В целом следует подчеркнуть, что экологические популяции не занимают определенного географического района, но в его условиях приурочены к разным типам местообитаний. Лишь часть экологических популяций вполне способна к самовоспроизведению, другие существуют либо благодаря притоку иммигрантов, либо временами полностью вымирают и восстанавливаются иммигрантами (Беклемишев, 1960). Разнообразие экологических популяций служит показателем экологической пластичности вида. Элементарные популяции. Элементарные популяции – обитатели отдельных стаций в неоднородных, комплексных биотопах. Это мелкие группы, заселяющие мозаичный ландшафт. Изоляция их друг от друга очень слаба и постоянно идет процесс обмена особями. Вообще наличие элементарных популяций и характерно только тем видам, или отдельным географическим или экологическим популяциям видов, чей характерный биотоп мозаичен. Элементарные популяции занимают в нем благоприятные места (стации переживания), а прочая территория (стации расселения) заселяется временно. В некоторых случаях часть особей может попадать и удерживаться в неблагоприятных местах долгое время, как полихета Nereis succinea в опресненных водах Каспия, где невозможно ее размножение. Такие поселения иногда называют псевдопопуляциями. У амфибийных животных разные стадии онтогенеза образуют свои популяции. В.Н.Беклемишев предложил называть их гемипопуляциями. Различная судьба отдельных элементарных популяций зависит от особенностей микро климата, состава кормов, убежищ и разного уровня гибели и размножения в этих стациях. Взаимоотношения между популяциями. Вид представляет собой единое образование, и его отдельные популяции связаны между собой. Связь популяций одного или разных рангов осуществляется с помощью перемещений животных, которые для существования вида также важны как размножение. С их помощью осуществляется встреча самцов с самками, устраняется опасность имбридинга, происходит перераспределение животных в зависимости от размещения кормов, микроклиматических условий. Наконец, происходит расселение животных в новые места и порой расширение ареала вида. Этому способствуют все виды миграций, в том числе периодические (сезонные, суточные) и непериодические (неурожай корма, засуха, наводнение). Особое значение играют миграции, связанные с размножением: перегруппировки в начале и конце сезона размножения и расселение молодняка. Несмотря на часто ярко выраженный гнездовой консерватизм, на зимовках популяции, хотя и редко, обмениваются особями. Часть видов совершают большие, но достаточно хаотичные миграции (песцы). Подвиды у этих видов выделить сложно. Другим видам свойственны правильные закономерные миграции (северные олени). Во время осенних кочевок многие воробьиные птицы входят в состав смешанных многовидовых стай, и здесь происходит обмен особями между популяциями. Важную роль в структуре вида играет расселение молодняка, которое свойственно даже неподвижным животным. У водных животных молодняк иногда преодолевает огромные пространства (пассивная миграция личинок речных угрей, скумбрии в Черном море, с ветром и самостоятельно некоторых насекомых – саранчи). Гнездовой консерватизм мелких воробьиных хорошо проявляется у взрослых и почти отсутствует у молодых птиц, которые широко расселяются в окрестностях мест кольцевания. Очень важен тот факт, что подвижность и большое количество мигрантов характерно не только на пике численности при переуплотнении популяции, но и при низкой численности, когда оно оказывается иногда даже более напряженным. Такой повышенной подвижностью животные поддерживают связи в редеющей популяции. Различие в подвижности разных половых и возрастных групп имеет большой биологический смысл. Расселение и потери среди молодых особей наносят меньший вред популяции в целом, малая подвижность в период размножения у самок также энергетически выгодна. Непериодические миграции и расселение молодняка на подъеме численности расширяют обычную область обитания вида и могут в определенных условиях привести к устойчивому расширению ареала. Подобные скачки границ известны для зайца-русака, леммингов и некоторых видов насекомых (стрекоз, саранчи, комаров). Путем выселения из оптимальных зон поддерживается заселение периферийной части ареала, где высокая гибель особей не покрывается размножением. Постепенно при этой подпитке на периферии идет приспособление вида к новым условиям. Периферические популяции, как правило, характеризуются рядом эколого-физиологических особенностей. Известно увеличение величины выводка у птиц и млекопитающих, удлинение суточной активности, рост индивидуальных участков, изменения в характере питания (увеличение его разнообразия). Массовые миграции, поддерживая периферию ареала, способствую приспособлению вида к новым условиям и расширению ареала. Кроме того, они необходимы для сохранения целостности вида. Из всего сказанного видно, что территориальные группировки, будучи самостоятельными, находятся в определенных отношениях друг с другом. Видовое население живет как единое целое, обладающее определенной исторически сложившейся пространственной структурой и организацией. Поддержание внутрипопуляционного гомеостаза связано, прежде всего, с такими показателями, как численность и плотность населения. Все динамические процессы в популяции основываются с одной стороны на общих для всех особей популяционных признаках, а с другой стороны на разнообразии особей по полу, возрасту, генетическим особенностям и месту в функционировании популяции. |