Скелет свободного крыла. Два звена основного столба конечности выражены полно, а дистальное звено сильно изменено и значительно редуци-. ровано.
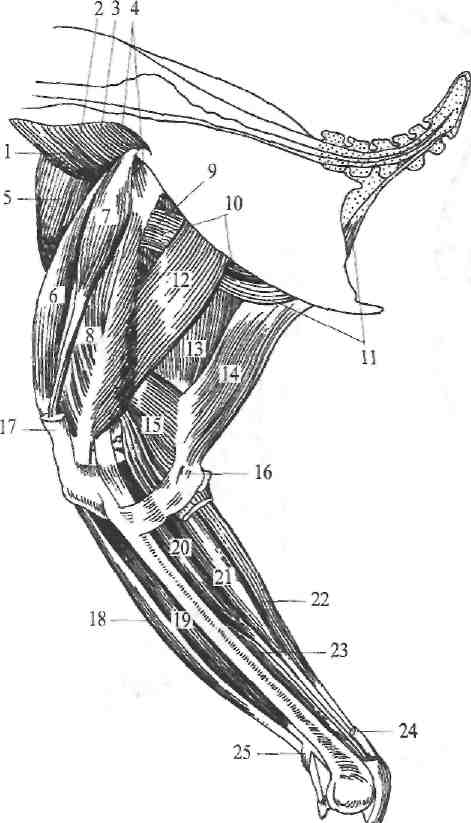
Плечевая кост ь—os humeri (33)—трубчатого типа, сильно развита и имеет утолщённые концы. При покое крыла она располагается на грудной клетке и направлена от плечевого сустава назад, к тазу. Её проксимальный (плечевой) конец в виде слабо выпуклой овальной головки соединяется с впадиной, образованной совместно лопаткой и коракоидом. Этот конец значительно утолщён, благодаря наличию на нём латерального (или малого). и медиального (или большого) бугорков. Здесь же с медиальной стороны имеется отверстие—foramen pneumaticum,—ведущее в воздухоносную полость, кости. Дистальный конец слегка утолщён и несёт на себе с волярной стороны полукруглую суставную поверхность для локтевой кости, а с дорзальной стороны—яйцевидную поверхность для лучевой кости.
Кости предплечь я—ossa antebrachii. В звене предплечья силь-. нее развита локтевая кость—ulna (35). Она слегка искривлена и снабжена, слабо выраженным локтевым отростком—olecranon. Почти прямая, более тонкая лучевая кость—radius (34)—лежит дорзо-медиально. Между обеими костями остаётся довольно широкое межкостное пространство—spatium interosseum. Предплечье при покое крыла расположено почти параллельно плечу, т. е. от локтевого сустава направлено вперёд.
Дистальное звено крыла, соответствующее передней лапе (кисти) ного-образных конечностей, весьма сильно изменено и приспособлено к односторонней функции полёта. Это изменение выразилось в редукции отдельных членов во всех участках этого звена. Само звено при покое крыла лежит под острым углом к предплечью и дистальным концом направлено назад и слегка опущено вниз.
3 а п я с т ь е—carpus (36, 37)—сохраняется только в виде запястной лучевой—os radiale—и запястной локтевой—os ulnare—костей; запястная промежуточная кость слита с запястной лучевой, а добавочная—с запястной локтевой. Дистальный ряд костей запястья полностью сливается с пястными коетями.
П я с т ь—carpometacarpus (56)—редуцирована до трёх члеников (2,3, 4-й), да и те слиты между собой в одно образование, к которому приращён и дистальный ряд запястья. Из них наиболее заметны 3 и 4-я пястные кости, скреплённые своими концами и имеющие между собой межкостное пространство. Вторая маленькая пястная косточка полностью слита с проксимальным концом соседней.
Скелет пальцев (39, 40, 41) также сильно редуцирован. Наиболее ясно сохранён 3-й палец с двумя фалангами; 2 и 4-й пальцы маленькие и, как правило, состоят из одной фаланги (о связи локтевого и карпаль-ного суставов см. раздел о мускулатуре).
918 СИСТЕМА ОРГАНОВ ПРОИЗВОЛЬНОГО ДВИЖЕНИЯ
Скелет тазовых конечностей
Тазовые конечности являются органами передвижения птиц по суше и воде. .
Тазовый пояс состоит из подвздошной, седалищной и лонной костей. У птиц он особенно характерен, во-первых, тем, что вентральные звенья (лонная и седалищная кости) правой и левой сторон не соединены друг с другом по сагиттальной линии, вследствие чего таз птиц широко открыт с нижней стороны. Во-вторых, таз птиц прочно и на большом протяжении слит с позвоночным столбом.
•Подвздошная кост ь—08 ilii (28)—сильно развита. Она ко-стно сращена с цояснично-крестцовым отделом позвоночника и простирается вперёд над позвоночными концами последних рёбер на грудной отдел.
Седалищная кост ь—os ischii (29)—служит как бы продолжением подвздошной кости назад и также сращена с позвоночным столбом. Подвздошными и седалищными костями, а также прилежащим участком позвоночника в совокупности создаётся обширная крыша тазовой полости. На вентральной поверхности этой крыши, у места сращения таза с поперечнорёберными отростками и телами позвонков, имеется ряд углублении (рис. 252), в большей части которых расположены ночки птиц. Между подвздошной и седалищной костями позади тазовой впадины находится седалищное отверстие— foramen ischiadicum (рис.250—31)—овальной формы.
Лонная кост ь—os pubis (30).—в форме длинной ленты лежит вентральнее седалищной кости. Её задний конец простирается за край седалищной кости и слегка изгибается медиально. Между седалищной и лонной костями помещается длинное запертое отверстие—foramen obturatum(-32). На месте сращения всех трёх костей таза сформирована тазовая впадина-— acetabulum—для сочленения с бедренной костью.
Свободная тазовая конечность, как и у других наземных позвоночных, состоит из трёх звеньев: бедра, голени и задней лапы (стопы).
Бедренная кост ь—os femoris (42)—короче костей голени. Её проксимальный конец имеет головку—caput femoris,—сочленяющуюся с тазом, и вертел—trochanter. От тазобедренного сустава бедренная кость опускается вниз и вперёд, причём её тело слегка искривлено в спинковую сторону. Дистальный конец снабжён мыщелками—condyli,—сочленяющимися с костями голени, а также блоком для коленной чашки—patella (43).
Кости голени. Болыпеберцовая кость—tibia (45)—большая и длинная. У птиц по особенному устроен её дистальный конец, так как с ним срастается в одно целое весь проксимальный ряд костей заплюсны. В силу такого слияния эту кость называют большеберцово-заплюсневой костью—tibiotarsus. Дистальный конец её несёт на себе две суставные поверхности наподобие мыщелков. Проксимальный конец сочленяется главным образом с медиальным мыщелком бедренной кости, так как латеральный её мыщелок сочленяется с малоберцовой костью. Малоберцовая кость—fibula (44)—сильно редуцирована и сращена с болыпеберцовой.
Скелет задней лапы (стопы) в своей структуре значительно упрощён по сравнению со скелетом того же звена других наземных четвероногих животных.
Кости заплюсны как самостоятельные элементы скакательного сустава отсутствуют. Проксимальный ряд сустава, как уже отмечено, слился с концом болыпеберцовой кости голени, а центральная кость и дистальный ряд костей заплюсны полностью срослись с костями плюсны. В силу сказанного в заплюсне сохранён лишь межзаплюсневый сустав.
Кости плюсн ы—metatarsus (46)—довольно длинные и представляют одно крепкое целое, получившееся в результате слияния главным об-ч
МУСКУЛАТУРА 919
р  азом трёх плюсневых костей (2,3 и 4-й). К ним проксимально приращены кости заплюсны, так что весь этот монолитный участок звена лапы называют заплюсно-плюсневой костью (цевкой)—tarsometatarsus. Вторая, третья и четвёртая плюсневые кости разделены только на дистальном конце, где все три кости несут суставные поверхности для сочленения с фалангами пальцев. На плантарной поверхности стопы, ближе к медиальному краю, имеется ещё маленькая первая плюсневая кость, сочленяющаяся с 1-м пальцем. Над ней у петухов развивается шпорный отросток. Скелет пальцев в наиболее полном виде насчитывает четыре луча: один задний— по счёту первый—и три передних—2, 3 и 4-й. Задний (1-й) палец (48) обладает двумя фалангами; из передних: 2-й—тремя, 3-й— четырьмя, 4-й—пятью фалангами. Следует, однако, отметить, что число пальцев, их величина и размещение у различных птиц сильно варьируют. МУСКУЛАТУРАУ птиц нелетающих или летающих с трудом (куры) мускулатура характеризуется бледным цветом, у остальных отрядов она тёмнокрасная. В сухожилиях мускулов часто встречаются окостеневшие участки.
1 2
Рис. 255. Мускулатура головы курицы.
А—п оверхностный слой: /—m. cuta-neus (обрезан); 2—т. temporalis; 3—го. masseter; 4—т. quadratomaxillaris; 5—т. digastricus; в—т. ту-lohyoideus; 7—т. geniohyoideus. В—с р е д н и й слой: 1—т. temporalis; 2—т. pterygoideus inter-nus; 3—-т. sphenomaxillaris; 4—т. quadratomaxil-laris; 5—т. levator palpebrae inferior!*. С—Г л у-бокий слой: /—т. pterygoideus externus;
MvPKVJTflTvna rnimnu TTwttp- 2m- pterygoideus intern us; 3—m. levator quadrati;
1У1 у О культура ГОЛОВЫ. ЛИЦе- 4—т. rectus oculi; 6, в—т. obliquus oculi superior
вая, или мимическая, мускулату- et inferior.
pa для движения губ, носа и
щёк отсутствует. Жевательные мускулы сильные и более диференцирован-ные, чем у млекопитающих, так как имеются отдельные мускулы, управляющие квадратной костью; они выдвигают её вперёд и оттягивают обратно,
Кожные мускулы птиц хорошо развиты и в большей степени распространены, чем у млекопитающих. Большинство их оканчивается на перьевых влагалищах показывает разностороннее действие на перья. Сокращением этих мускулов расправляются при полёте маховые и рулевые перья. Другие кожные мускулы напрягают летательную перепонку крыла (mm. patagii s. propatagiales s. mm. plicae alaris (рис. 258— 24; 259— 11, 12). Кожные мускулы встречаются не только на туловище и шее, но и в области затылка и лба (рис. 255— 1).
920
СИСТЕМА ОРГАНОВ ПРОИЗВОЛЬНОГО ДВИЖЕНИЯ
причём это движение обусловливает соответственно приподнимание и опускание надклювья, передаваясь последнему через цепь из крыловидных и нёбных костей, а также через скуловую дугу (см. «Скелет головы»). 5048  К  комплексу жевательных мускулов (рис. 255) принадлежат: m. masseter (А—3), т. pterygoi-deus externus et interims (В, 2; С, 1), т. temporalis (А, 2), т. quadratomaxillaris (A, 4), m.sphenomaxillaris (В, 3), т. levator quadrati (С, «3),т. digastricus (А, 5). Ряд мускулов управляет Рис. 256. • Мускулатура курицы. 8г"\ 9—т. splenius capitis; \. 10—га. rectus capitis ^-55 lateralis; 11—т. biventer cervicis; 12—m. tracheo- mastoideus; 73—m. rectus capitis ventralis maior; 14—ram. intertransversa- rii; 16—m. flexor colli brevis; j6—m.
sternotrachealis; 17—m. sternohyoideus; 18—m. extensor cervicis Iongus; 19—m. cervicalis descenders; 20—m. spina lis dorsi et cervicis; 21—m. longus colli; -72 26—m. anconeus longus; 48—m. latis-
-81
72-
^Г
simus dorsi; 60—mm. plicae alaris; 61—m. rhomboideus; 52—m. suprasca- pularis; 63—m. serratus centralis; 64—m. obliquus abdominis externus; 66—m. pectoralis maior; 66—m. sar- torius; 67—m. tensor fasciae latae; 68—m. biceps femoris (caput longum); 59—m. semitendinosus; 60—m. obli quus abdominis externus; 61—m. bi ceps femoris (caput breve); 62—m. le vator ani; 63—m. sphincter ani; 64—m. pubococcygeus; 65—m. ischioeoccygeus; 67, 68—mm. levatores caudae; 69,
70 — m. gastrocnemius (caput mediale et cap. laterale); 71—m. flexor digiti tertii longus; 72—m. soleus et peroneus longus; 78—m. flexor hal- lucis longus; 79—m. fle xor digitalis profundus; 81—m. abductor digiti quarti; b — трахея; с—аоб; d—копчиковая железа; е—копчик. подъязычной костью и тем самым приводит в движение язык, который имеет в своей толще весьма слабо развитые мускулы. Мускулатура туловища. Дорзальные мускулы грудного и пояснично-крестцового отделов позвоночного столба очень слабые (рис. 257 — 5; 261— 5), так как эти отделы или незначительно, или совершенно неподвижны (см. описание скелета позвоночного столба). Довольно богата мускулатура, идущая к хвостовому отделу позвоночника (рис. 261 — 16, 16, 17, 18).
МУСКУЛАТУРА 921
Благодаря её наличию хвостовый отдел приобретает разностороннюю подвижность.
Брюшные мускулы имеются, как у млекопитающих, но представляют очень тонкие и слабые пласты. К ним относятся: m. obliquus abdominis externus (рис. 256—5/, 60; 2.57—17), т. obliquus abdominis inter-nus {22), т. transversus abdominis и т. rectus abdominis (21).
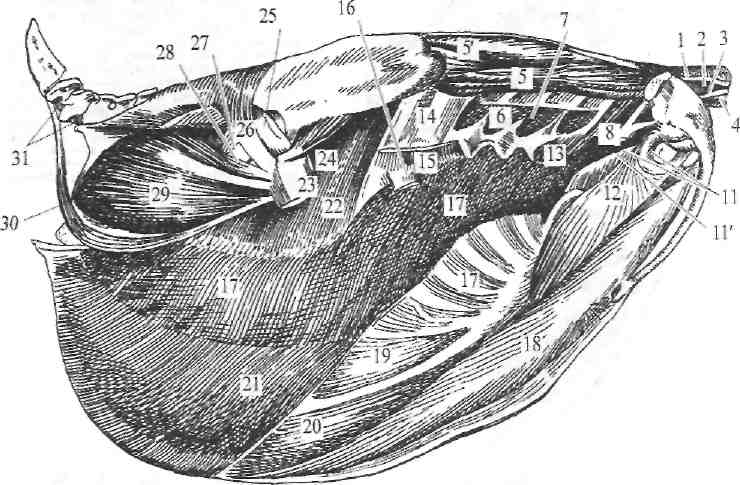
Рис. 257. Мускулатура наружной стороны туловища курицы.
1—m. biventer; 2—т. extensor colli longus; 3—т. cervicalis descendens; 4—т. spinalis dorsi; 5—т. sacrolumbulis; s'—m. longissimus dorsi; 6—mm. levatores costarum; 7—m. levator scapulae; 8—m. suprascapularis; 11, 11'—m. coracobrachialis; 12—m, pectoralis tcrtius; 13—m. ser-ratus ventralis minor; 14—mm. intercostales; 15—m. serratus ventralis maior; 16—m. plicae alaris posterior; 17, 17 —m. obliquus abdominis externus; 18—m. pectoralis minor; 19, 20—мембраны между отростками грудины; 21—m. rectus abdominis виден иод сухожилием m. obliquus abdomi is externus; 22—m. obliquus abdominis internus виден иод сухожилием m. obi. abd. ext.; 23—m. glutaeus medins; 24—m. glutaeus profundus; 25—m. glutaeus superficialis; 2G—m. piriformis; 27—m. obturator internus; 28—m. quadratus femoris; 29, 30—mm. gemelli;
31—m. depressor caudae.
Мускулатура грудной клетки включает: mm. intercostales externi (14), mm. intercostales interni, mm. levatores costarum (6). m- transversus thoracis и mm. scaleni. Своим действием все они оказывают влияние па движение грудной клетки при дыхании. Акт вдыхания выражается главным образом разведением углов между вертебральными п стернальны-ми участками рёбер, сопровождаемым опусканием грудной кости (рис. 278).
Диафрагма имеется, но она представляет сухожильную пластинку, расправленную по вентральней поверхности лёгких. Её весьма слабые мускульные зубцы подходят к ней от рёбер.
Шейная мускулатура сильно развита и диференцирована, особенно в участке, ближайшем к голове (рис. 256). Вся эта мускулатура делает возможньш самые разнохарактерные движения шеи и головы около шеи.
Мускулатура крыла состоит из довольно большого числа мускульным единиц (рис. 256, 257, 258 и 259). Среди них особенно выделяются сильные грудные мускулы—mm. pectcrales maior, tertius, minor (рис. 256—55\ 257—12, 18).
922
СИСТЕМА ОРГАНОВ ПРОИЗВОЛЬНОГО ДВИЖЕНИЯ
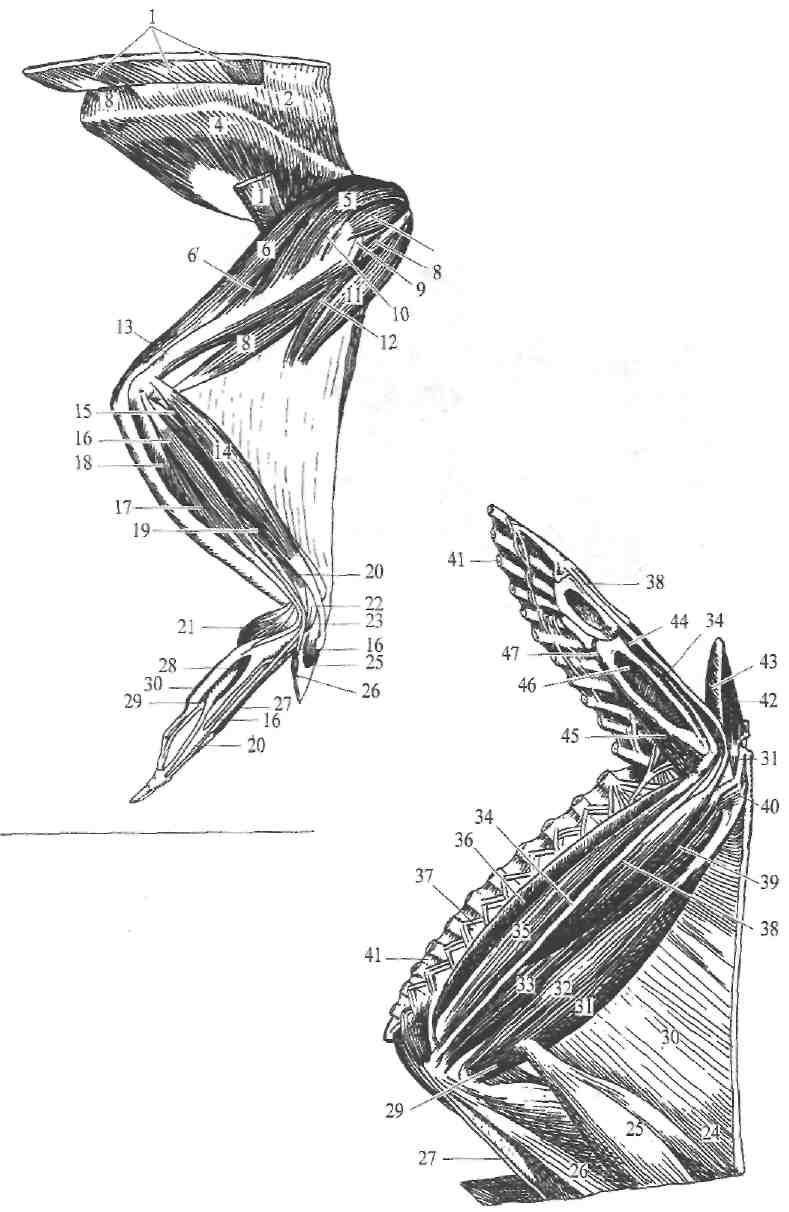
Рис. 259. Мускулатура крыла курицы с латеральной стороны. 1—m. latissiraus dorsi; 2—т. trapezius; 3—т. rhomboideus; 4—т. suprascapu-laris; 5—т. deltojdeus maior; 6—т. anconeus longus; в'—т. anconeus interims; 7—т. deltoideus minor; *—m. biceps brachii; 9—m. deltoideus medius; 10—ш. pectoralis minor; 77, 12—m. plicae alaris; 13—m. anconeus minor; 14—m. extensor carpi radialis longus; 16—m. supinator; 16—m. extensor digitalis brevis; 17—m. extensor carpi ulnaris; is—m. flexor antebrachii profundus; 7.9—m. extensor carpi radialis brevis; 20—m. extensor digitalis longus; 21—m, flexor raeta-carpi brevis; 22—m. extensor indicis; 23—сухожилие от m. fiexoris carpi radialis; 25—та. extensor pollicis longus; 26—m. flexor pollicis; 27—m. abductor iodicis; 28—m. adductor indicis; 29—m. interosseus interims; 30—m. flexor digiti minimi.
Ряс. 258. Мускулатура крыла
курицы с медиальной стороны.
24—m. plicae alaris; 25—т. biceps brachii; 26—т. anconeus longus; 27—сухожилие от т. coracobrachialis; 20—га. brachialis; 30—plica alaris; 31—га. extensor carpi radialis longus; 32—m. pronator longus; 33—m. pronator brevis; 34—m. flexor digitalis sublimiS; 35—in. flexor carpi ulnaris; 36—m. palmarls longus; 31—мускулы перьев ;38 — т. flexor digital is profundus; 39— m. extensor digitalis longus; 40—m. flexor carpi radialis; 41—перья (обрезаны); 42—т. extensor pollicis longus; 43—m. flexor pollicis; 44—m. abductor indicis; 45—m. flexor metacarpi brevis; 46—m. interosseus internuS; 47—m. flexor digiti minimi.
МУСКУЛАТУРА 923
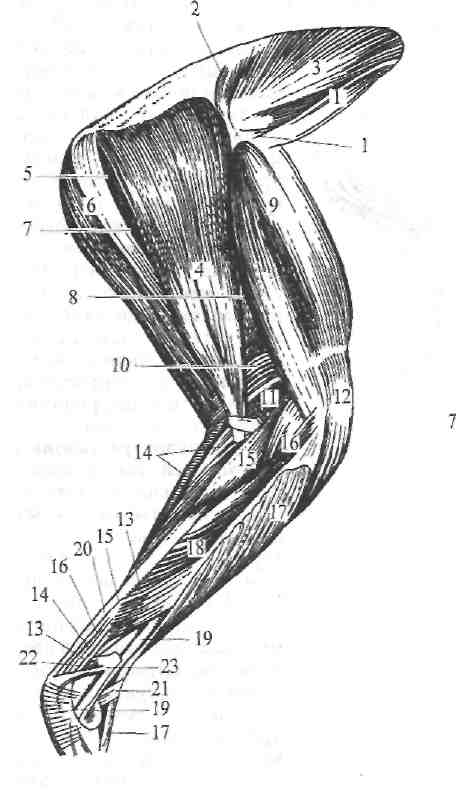
Рис. 260. Глубокая мускулатура наружной поверхности тазовой конечности курицы.
/, 3—mm. glutaei; 2—m. piriformis; 4—m. biceps femoris (caput breve); 5—m. gemellus; в—m. semitendinosus; 7—m. crurococcygeus; 8—m. adductor longus; 9-—m. quadriceps femoris; 10, 11, 12—m. m. gastrocnemii; 14—n. Ilexor digiti quarti longus; 15—m. flexor digiti secundi longus; 16—m. flexor digiti tertii longus; 17—m. tibialis anterior; IS—m. flexor digitalis profundus; 19—m. peroneus tertius; 20—сухожилие mm, gastrocnemii; 21—связки; 22—m. flexor hallueis longus; 23—m. soleis et peroneus longus (обрезан).
234
-
'
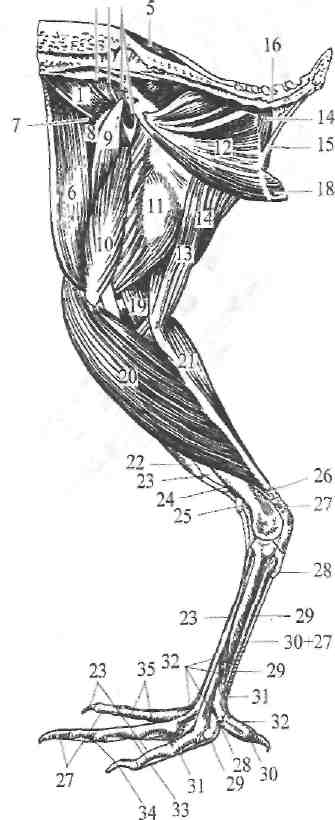
Рис 261. Мускулатура тазовой
конечности курипы с медиальной
стороны.
1, 2, 3—mm. glutaei; 4—т. iliacus internus; 5—m. semispinals dorsi; 6—m. sartorius, 7—m. tensor fasciae liatae; 8—m. rectue femoris; 9—m. gracilis; 10—m. vastus medialis; 11—m. adductor magnus; 12—obturator internus; 13—m. semimembranosus; 14—m. semitendinosus; IS—m. crurococcygeus; IS—m. depressor caudae; 17—m.pubo-coccygeus; 18—m. ischlococcygeus; 19, 20, 21—mm. gastrocnemii; 22—m. soleus et peroneus longus;23—m. extensor digitalis longus; 24—m. tibialis anterior; 25—связки; 26—m. tibialis posterior; 27—m. flexor digitalis profundus; 28—m. flexor hallueis brevis; 29—m. flexor digiti secundi brevis; 30—m. flexor hallueis brevis; 31—m. flexor digiti seaundi longus; 32—m. extensor digitalis brevis; 33—m. flexor digiti tertii brevis; 34—m. flexor digiti tertii longus; 35—-m. flexor digiti quarti longus.
924
СИСТЕМА ОРГАНОВ ПРОИЗВОЛЬНОГО ДВИЖЕНИЯ
Следует отметить сочетанную функцию локтевого и запястного суставов, выражающуюся в том, что при разгибании локтевого сустава разгибается и запястный сустав. То же самое имеет место и в процессе сгибания. Эта сочетанность действия обусловливается тем, что локтевая и лучевая кости лежат в предплечье своими концами на разных уровнях (локтевая кость несколько выше лучевой), причём концы их имеют с. соседними звеньями отдельные сочленения. Отсюда понятно, что мускулы, действующие на локтевой сустав, обладают сравнительно большой мощностью; их сокращение отражается и на функции запястного сустава. Мускулатура тазовых конечностей также включает довольно значительное количество отдельных единиц, большинство которых соответствует таковым млекопитающих (рис. 256, 257, 260, 261, 262).
Рис. 262. Глубокая мускулатура тазовой конечности курицы с медиальной стороны.
1,2, 3—mm. glutaei; 4—m. iliacus internus; 5—m. tensor fasciae latae; в—m. rectus femoris; 7—m. gracilis; 8—m. vastus medians; a—m. quadratus femoris; 10,11—mm. gemeili; 12—m. adductor longus; 13—m. biceps famoris (caput breve); 14,—m. se-mitendinosus; 15—m. gastrocnemius; 16 — m. semimembranosus (обрезан); 17—m. sartorius (обрезан); 18—m. tibialis anterior; 19—m. extensor digitalis longus; 20—m. tibialis posterior; 21—m. flexor digiti tertii brevis; 22—m. flexor digiti quarti longus; 23—m. flexor digitalis profundus; t*—сухожилие m. gastrocnemii; 25—связки.
Особенно массивны мускульные брюшки в звене бедра. Отдельного упоминания заслуживает специальное сухожильное приспособление, оказывающее своё действие на пальцы. Это сухожилие ответвляется от стройного мускула, тянется по сшга-ковой поверхности коленной чашки, затем переходит через латеральную сторону скакательного су • става на плантарную поверхность голени и сливается с поверхностным пальцевым сгибателем. На своём пути сухожилие прикрепляется к проксимальному концу малоберцовой кости. Благодаря такому положению сухожилия при сгибании коленного сустава одновременно механически сгибаются фаланги пальцев. Это простое приспособление вступает в действие при различных состояниях конечностей птиц. Когда птица садится на сучок и тяжестью тела сгибает коленный сустав, её пальцы, механически сгибаясь, крепко удерживают сучок, так что
СИСТЕМА ОРГАНОВ КОЖНОГО ПОКРОВА 925
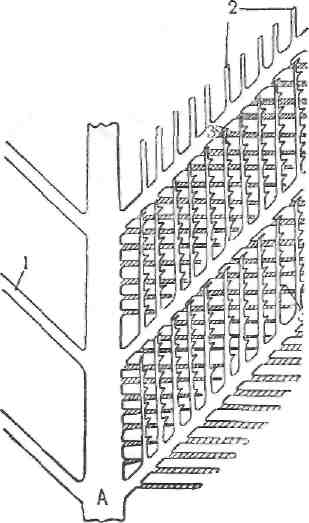
птица может спать в сидячем положении, не затрачивая мускульной силы. При движении по суше этот механизм помогает действию флексоров пальцев в фазе переноса ноги на новое место. При этом характерно, что сгибание пальцев сопровождается схождением их лучей, а разгибание— расправлением. Это происходит благодаря особому устройству суставных поверхностей дистального конца плюсневых костей. II. СИСТЕМА ОРГАНОВ КОЖНОГО ПОКРОВА Птицам присущи обычные напластования кожного покрова: а) надкожица, б) основа кожи и в) подкожный слой. Надкожица—epidermis—на местах, густо покрытых перьями, очень тонкая, и поверхностный слой её постоянно отделяется—шелушится. На клюве, когтях, мякишах пальцев, че-шуях ног и шпорах петуха роговой слой сильно развит; основа кожи в таких участках содержит ясно выраженный сосочковый слой. Основа кожи—corium —также сравнительно тонка, безжелезиста и не отличается обилием сосудов; только на таких образованиях головы, как гребень и бородки, сосудами формируется богатая сеть, напоминающая структуру кавернозного тела. Подкожный слой—subcutis—тонкий и всюду ясно выражен; он содержит в ряде мест подкожные синовиальные сумки и жировую ткань. Одной из характерных особенностей строения системы кожного покрова птиц служит отсутствие в ней (как у рептилий) потовых и сальных желез. Только над последними крестцовыми позвонками у большинства птиц находится так называемая копчиковая железа—glandula uro-pygii (рис. 256— d). Она состоит из двух различной величины долей, причём се-кретирующие отделы выстланы много- Рис- 263- Схема строения опахала слойным эпителием, функционирующим А_ стебель (rachis). 7_ ветви (rami) наподобие сальной железы. Выводные г—лучи (radii); з—крючки (hamuli), протоки от секретирующих отделов открываются в небольшую полость, от которой наружу ведут один или два (редко больше) хода. Они открываются рядом с таковыми другой доли на особом возвышении кожного покрова. Секрет копчиковой железы содержит жир и служит для смазывания перьев, поэтому у водоплавающих птиц железа развита сильнее (у кур её доли величиной с горошину, а у уток—с лесной орех). Смазывание перьев происходит путём переноса клювом секрета железы. Перья. Второй, ещё более яркой особенностью покрова птиц служит оперение. Оно имеет огромное значение для птиц не только при полёте, но и способствовало на ранних этапах эволюции, как и волосы млекопитающих, установлению и удержанию постоянства температуры тела. На развитом пере различают: с т е р ж е н ь—scapus—и опахал о— vexillum (или бородку—barba). В свою очередь, стержень подразделяется
926
СИСТЕМА ОРГАНОВ КОЖНОГО ПОКРОВА
на о чи н—calamus—и стебел ь—rachis. Очин представляет начальную, свободную от опахала часть пера, сидящую главным образом в перьевой сумке толщи кожи, а стебель—остальную стержневую часть пера. Опахало построено довольно сложно и состоит из ветвей—rami (рис. 263— 1), отходящих от стебля в две противоположные стороны. От ветвей, в свою очередь, отделяются лучи—radii (2). Лучи снабжены крючками—hamuli (s. radioli) (3). Возникновение пера в начальной стадии весьма схоже с появлением у рептилий чешуи, которые впоследствии преобразовались в перья. 
Рис. 264. Схема развития пера.
А—начальная стадия: 1—эпителий пера; 2—мезенхима (зачаток сосочка); 3—кожа. В—дальнейшая стадия с образованием перьевой сумки: 1—перьевая сумка; 2—соединительнотканный сосочек (пульпа пера); 3—эпителий опахала; 4—кожа. С—поперечный разрез зачатка опахала пушкового пера; 7—пульпа пера; 8—-формирующиеся из эпителия стебельки. О—то же самое на дальнейшей стадии; 9—отъединившиеся друг от друга стебельки опахала; 10—соединительнотканная часть пера. Е—готовое молодое пушковое перо; 4—стебельки; 6—полость перьевой сумки; 6—пульпа очина; 7—окружающая кожа. F—формообразование опахала контурных, рулевых и маховых перьев; 11—стебель опахала; 12—перьевая сумка; 13—сосочек пера; 14—ветви и лучи опахала; 15—очин. G—вырезанный участок развивающегося опахала: 16, 17—стебель; 18, 19—ветви, отходящие от стебля 20—сходящиеся вершины ветвей.
В местах эмбрионального развития перьев вначале наблюдается скопление мезенхимных клеток непосредственно под эпидермисом, который приподнимается с отклонением назад по отношению к оси тела (рис. 264—А,1). С дальнейшим ростом этого зачатка окружающий его участок кожи углубляется, формируя закладку перьевой сумки, или влагалища (В,1).
Уже в этом простом зачатке, помимо влагалища, намечены лежащий центрально и богатый сосудами соединительнотканный сосочек (пульпа пера) (В, 2) и окружающий его эпителий (3). Последний распределяется в два слоя—поверхностный и глубокий. Первый состоит из плоских клеток и формирует временный чехол для развиваюшегося молодого пера. Глубокий цилиндрический слой эпителия, разрастаясь, даёт начало отдельным
СИСТЕМА ОРГАНОВ КОЖНОГО ПОКРОВА
927
частям пера. Ранний процесс формообразования завершается появлением эмбрионального пушка. Клетки эпителия под чехлом образуют приблизительно одинаковые продольные выросты внутрь пера к его пульпе (С, 8). Эти выросты, ороговевая, отъединяются от чехла и друг от друга на выступающем участке пушкового пера (D, 9) и становятся отдельными элементами (стебельками) опахала. По сбрасывании чехлика они разъединяются окончательно, и пушок готов (Е, 4).■  5 Рис. 265. Крыло. 1, 2, 3—контурные перья; 4— маховые перья; 5—крылышко.
Рис. 266. Распределение покровных перьев
А—вид с вентральной стороны; В—вид с дор-
зальной стороны.
Перьевые (pterylae) и бесперьевые (apteriae) облает» кожного покрова курицы. Pterylae заштрихованы.
Э  тот пушок в дальнейшем сменяется перьями взрослых, причём последние возникают под пушком на том же самом сосочке и вытесняют первое поколение. Развитие дефинитивного пера протекает с некоторым усложнением, которое выражается в том, что разроет эпителия под чехлом внутрь совершается неравномерно и не строго вдоль пера. Один из отпрысков оказывается очень массивным и превращается в зачаток стебля— rachis (F,ll; G, 16). Остальные выросты значительно меньше него; они дают ветви—rami (14, 19)—и располагаются по окружности пульпы под углом к стеблю (17) с двух сторон, сходясь вершинами (20) друг с другом. От них отделяются отростки—будущие лучи—radii (рис. 263— 2),—а затем и крючки— hamuli (3). По сбрасывании чехла ветви расправляются, формируя опахало. В результате этой наиболее сложной формы развития на коже появляются контурные, или покровные, перь я—реппае (рис. 265— 1, 2, 3). Если процесс развития останавливается до формирования крючков, то ветви опахала, сидящие на небольшом стебле, не сцепляются друг с другом и получаются пуховые перь я—plumae. Они меньше по размеру и лежат под покровными перьями, играя роль тёплого одеяния. Покровные перья развиваются на определённых участках кожного покрова, называемых pte-rylae, а промежутки между ними остаются без перьев и называются ар-teriae (рис. 266). На крыльях птиц находятся очень длинные, с широким опахалом, воздухоупорные маховые перь я—remiges (рис. 265— 4),—играющие |
 Скачать 1.33 Mb.
Скачать 1.33 Mb.