Ответы на билеты биохимия 2012. Белок это последовательность ак, связанных друг с другом пептидными связями
 Скачать 5.15 Mb. Скачать 5.15 Mb.
|

|
Р 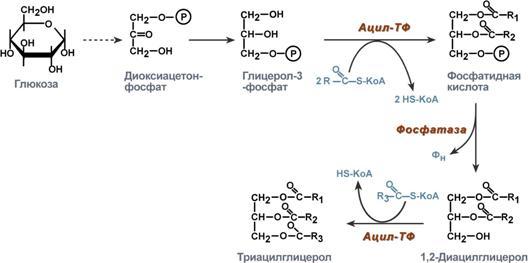 есинтез фосфолипидов. Фосфолипиды синтезируются также как и в остальных клетках организма. Для этого есть два способа. Первый – с использованием 1,2-ДАГ и активных форм холина и этаноламина для синтеза фосфатидилхолина или фосфатидилэтаноламина. Второй путь – через синтезируемую in situ фосфатидную кислоту. После ресинтеза фосфолипиды, триацилглицеролы, холестерол и его эфиры упаковываются в особые транспортные формы липидов – липопротеины и только в такой форме они способны покинуть энтероцит. В кишечнике формируются два вида липопротеинов – хиломикроны и липопротеины высокой плотности. есинтез фосфолипидов. Фосфолипиды синтезируются также как и в остальных клетках организма. Для этого есть два способа. Первый – с использованием 1,2-ДАГ и активных форм холина и этаноламина для синтеза фосфатидилхолина или фосфатидилэтаноламина. Второй путь – через синтезируемую in situ фосфатидную кислоту. После ресинтеза фосфолипиды, триацилглицеролы, холестерол и его эфиры упаковываются в особые транспортные формы липидов – липопротеины и только в такой форме они способны покинуть энтероцит. В кишечнике формируются два вида липопротеинов – хиломикроны и липопротеины высокой плотности. П 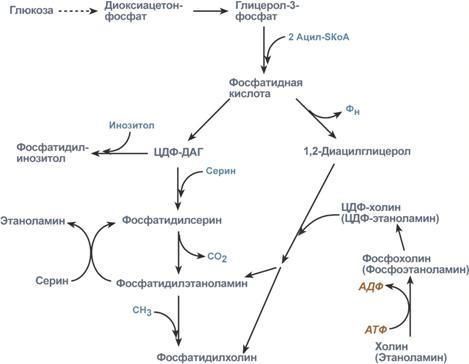 оскольку липиды являются в основе своей гидрофобными молекулами, то они транспортируются в водной фазе крови в составе особых частиц – липопротеинов. Такие транспортные липопротеины можно сравнить с орехом, который имеет скорлупу и ядро. Поверхность липопротеиновой частицы ("скорлупа") гидрофильна и сформирована белками, фосфолипидами и свободным холестеролом. Триацилглицеролы и эфиры холестерола составляют гидрофобное ядро. Липопротеины различаются по соотношению триацилглицеролов, холестерола и его эфиров, фосфолипидов и как сложные белки состоят из четырех классов.1. хиломикроны (ХМ), 2.липопротеины очень низкой плотности (ЛПОНП, пре-β-липопротеины, пре-β-ЛП), 3. липопротеины низкой плотности (ЛПНП, β-липопротеины, β-ЛП), 4. липопротеины высокой плотности (ЛПВП, α-липопротеины, α-ЛП). Транспорт триацилглицеролов от кишечника к тканям (экзогенные ТАГ) осуществляется в виде хиломикронов (ХМ), от печени к тканям (эндогенные ТАГ) – в виде липопротеинов очень низкой плотности. В транспорте ТАГ к тканям можно выделить последовательность следующих событий: оскольку липиды являются в основе своей гидрофобными молекулами, то они транспортируются в водной фазе крови в составе особых частиц – липопротеинов. Такие транспортные липопротеины можно сравнить с орехом, который имеет скорлупу и ядро. Поверхность липопротеиновой частицы ("скорлупа") гидрофильна и сформирована белками, фосфолипидами и свободным холестеролом. Триацилглицеролы и эфиры холестерола составляют гидрофобное ядро. Липопротеины различаются по соотношению триацилглицеролов, холестерола и его эфиров, фосфолипидов и как сложные белки состоят из четырех классов.1. хиломикроны (ХМ), 2.липопротеины очень низкой плотности (ЛПОНП, пре-β-липопротеины, пре-β-ЛП), 3. липопротеины низкой плотности (ЛПНП, β-липопротеины, β-ЛП), 4. липопротеины высокой плотности (ЛПВП, α-липопротеины, α-ЛП). Транспорт триацилглицеролов от кишечника к тканям (экзогенные ТАГ) осуществляется в виде хиломикронов (ХМ), от печени к тканям (эндогенные ТАГ) – в виде липопротеинов очень низкой плотности. В транспорте ТАГ к тканям можно выделить последовательность следующих событий:1)Образование незрелых первичных ХМ в кишечнике. 2)Движение первичных ХМ через лимфатические протоки в кровь. 3)Созревание ХМ в плазме крови – получение белков апоС-II и апоЕ от ЛПВП. 4)Взаимодействие с липопротеинлипазой (ЛПЛ) эндотелия, которая отщепляет жирные кислоты от ТАГ. Жирные кислоты переходят непосредственно в данную ткань или, связываясь с альбумином, разносятся по организму. В результате количество ТАГ в хиломикроне резко снижается и образуются остаточные ХМ. 5)Переход остаточных ХМ в гепатоциты и полный распад их структуры. 6)Синтез ТАГ в печени из пищевой глюкозы. Использование ТАГ, пришедших в составе остаточных ХМ. 7)Образование первичных ЛПОНП в печени. 8)Созревание ЛПОНП в плазме крови – получение белков апоС-II и апоЕ от ЛПВП. 9)Взаимодействие с липопротеинлипазой эндотелия и потеря большей части ТАГ. Образование остаточных ЛПОНП (по-другому липопротеины промежуточной плотности, ЛППП). 10)Остаточные ЛПОНП переходят в гепатоциты и полностью распадаются, либо остаются в плазме крови. После воздействия на них печеночной ТАГ-липазы в синусоидах печени ЛПОНП превращаются в ЛПНП. Характеристика хиломикронов. Общая характеристика: -формируются в кишечнике из ресинтезированных жиров, -в их составе преобладают ТАГ, мало белка, фосфолипидов и холестерола (2% белка, 87% ТАГ, 2% ХС, 5% эфиров ХС, 4% фосфолипидов), - основным апобелком является апоВ-48, это структурный липопротеин, в плазме крови получают от ЛПВП белки апоС-II и апоЕ, - в норме натощак не обнаруживаются, в крови появляются после приема пищи, поступая из лимфы через грудной лимфатический проток, и полностью исчезают через 10-12 часов,- не атерогенны. Функция: Транспорт экзогенных ТАГ из кишечника в ткани, запасающие или использующие жиры, в основном жировую ткань, легкие, печень, миокард, лактирующую молочную железу, костный мозг, почки, селезенку, макрофаги. На эндотелии капилляров этих тканей имеется фермент липопротеинлипаза. М 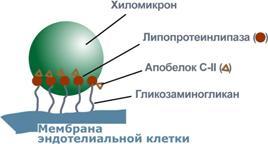 етаболизм 1. После ресинтеза жиров в эпителиоцитах кишечника формируются первичные хиломикроны, имеющие только апоВ-48. етаболизм 1. После ресинтеза жиров в эпителиоцитах кишечника формируются первичные хиломикроны, имеющие только апоВ-48.2. Из-за большого размера они не проникают напрямую в кровеносное русло и эвакуируются через лимфатическую систему, попадая в кровь через грудной лимфатический проток. 3. В крови хиломикроны взаимодействуют с ЛПВП и приобретают от них апоС-II и апоЕ, образуя зрелые формы. Белок апоС-II является активатором фермента липопротеинлипазы, белок апоЕ необходим для удаления из крови остаточных хиломикронов. 4. На эндотелии капилляров вышеперечисленных тканей находится фермент липопротеинлипаза (ЛПЛ), отщепляющий жирные кислоты от ТАГ. Количество фермента увеличивается при действии инсулина и прогестерона. 5. После взаимодействия хиломикрона с ферментом триацилглицеролы, находящиеся в составе хиломикронов, гидролизуются с образованием свободных жирных кислот. Жирные кислоты перемещаются в клетки органа, либо остаются в плазме крови и в комплексе с альбумином разносятся с кровью в другие ткани. Липопротеинлипаза способна удалить до 90% всех ТАГ, находящихся в хиломикроне или ЛПОНП. 6. После окончания работы ЛПЛ остаточные хиломикроны попадают в гепатоциты посредством апоЕ-рецепторного эндоцитоза и разрушаются. Содержание липидов в кровотоке может понижаться вслед¬ствие откладывания их в различных тканях. Способность откладывать жир характерна для всех тканей, кроме мозга. Главную роль в обмене липидов играют жировая ткань и печень. Количество жиро¬вой ткани нарастает с возрастом. Пределы конц жиров в крови:Норма-4-8г/л, если больше-гиперлипемия( при сах диабете, ожирении). В норме концентрация жиров в крови колеблется в довольно широких пределах — 10-200 мг/дл, в среднем около 0,1 %. Отметим для сравнения, что концентрация жиров в молоке равна примерно 3 %. После приема пищи концентрация хиломикронов в крови повышается, достигает максимума примерно через 5 ч, затем начинает снижаться. Сходным образом, но с меньшей амплитудой изменяется концентрация ЛОНП в крови. При этом надо отметить, что концентрация хиломикронов в большей мере зависит от содержания жиров в пище, а концентрация ЛОНП — от содержания углеводов. В крови, взятой для анализа утром до завтрака, т. е. после большого ночного перерыва в приеме пищи, хиломикроны не обнаруживаются, а концентрация ЛОНП минимальна. Пропорционально концентрации этих липопротеинов снижена и концентрация жиров в крови.При обычном ритме питания и небольших физических нагрузках в крови в дневное время постоянно имеются хиломикроны и ЛОНП, поскольку время переваривания жиров мало отличается от времени между приемами пищи. Любое нарушение внешнего обмена липидов (проблемы переваривания или всасывания) проявляется увеличением содержания жира в кале – развивается стеаторея. Причины: 1. Снижение желчеобразования в результате недостаточного синтеза желчных кислот и фосфолипидов при болезнях печени, гиповитаминозах. 2. Снижение желчевыделения (обтурационная желтуха, билиарный цирроз, желчнокаменная болезнь). У детей часто причиной может быть перегиб желчного пузыря, который сохраняется и во взрослом состоянии. 3. Снижение переваривания при недостатке панкреатической липазы, который возникает при заболеваниях поджелудочной железы (острый и хронический панкреатит, острый некроз, склероз). Может возникать относительная недостаточность фермента при сниженном выделении желчи. 4. Избыток в пище катионов кальция и магния, которые связывают жирные кислоты, переводят их в нерастворимое состояние и препятствуют их всасыванию. Эти ионы также связывают желчные кислоты, нарушая их работу. 5. Снижение всасывания при повреждении стенки кишечника токсинами, антибиотиками (неомицин, хлортетрациклин). 6. Недостаточность синтеза пищеварительных ферментов и ферментов ресинтеза липидов в энтероцитах при белковой и витаминной недостаточности. 22. Окисление глицерина и высших жирных к-т. Последовательность реакций. Связь β-окисления с циклом Кребса и дых цепью. Физиологическое значение окисления жирных кислот в зависимости от ритма питания и мышечной активности. Д 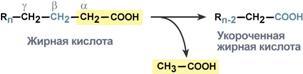 ля преобразования энергии, заключенной в жирных кислотах, в энергию связей АТФ существует метаболический путь окисления жирных кислот до СО2 и воды, тесно связанный с циклом трикарбоновых кислот и дыхательной цепью. Этот путь называется β-окисление, т.к. происходит окисление 3-го углеродного атома жирной кислоты (β-положение) в карбоксильную группу, одновременно от кислоты отщепляется ацетильная группа, включающая С1 и С2 исходной жирной кислоты. Реакции β-окисления происходят в митохондриях большинства клеток организма (кроме нервных клеток). Для окисления используются жирные кислоты, поступающие в цитозоль из крови или появляющиеся при липолизе собственных внутриклеточных ТАГ. Суммарное уравнение окисления пальмитиновой кислоты выглядит следующим образом: Пальмитоил-SКоА + 7ФАД + 7НАД+ + 7Н2O + 7HS-KoA → 8Ацетил-SКоА + 7ФАДН2 + 7НАДН ля преобразования энергии, заключенной в жирных кислотах, в энергию связей АТФ существует метаболический путь окисления жирных кислот до СО2 и воды, тесно связанный с циклом трикарбоновых кислот и дыхательной цепью. Этот путь называется β-окисление, т.к. происходит окисление 3-го углеродного атома жирной кислоты (β-положение) в карбоксильную группу, одновременно от кислоты отщепляется ацетильная группа, включающая С1 и С2 исходной жирной кислоты. Реакции β-окисления происходят в митохондриях большинства клеток организма (кроме нервных клеток). Для окисления используются жирные кислоты, поступающие в цитозоль из крови или появляющиеся при липолизе собственных внутриклеточных ТАГ. Суммарное уравнение окисления пальмитиновой кислоты выглядит следующим образом: Пальмитоил-SКоА + 7ФАД + 7НАД+ + 7Н2O + 7HS-KoA → 8Ацетил-SКоА + 7ФАДН2 + 7НАДН Э 2  . Ацил-S-КоА не способен проходить через митохондриальную мембрану, поэтому существует способ его переноса в комплексе с витаминоподобным веществом карнитином. На наружной мембране митохондрий имеется фермент карнитин-ацилтрансфераза I. . Ацил-S-КоА не способен проходить через митохондриальную мембрану, поэтому существует способ его переноса в комплексе с витаминоподобным веществом карнитином. На наружной мембране митохондрий имеется фермент карнитин-ацилтрансфераза I. Карнитин синтезируется в печени и почках и затем транспортируется в остальные органы. Во внутриутробном периоде и в первые годы жизни значение карнитина для организма чрезвычайно важно. Энергообеспечение нервной системы детского организма и, в частности, головного мозга осуществляется за счет 2х параллельных процессов: карнитин-зависимого ок-ия жирных к-т и аэробного окисления глюкозы. Карнитин необходим для роста головного и спинного мозга, для вз-вия всех отделов НС, ответственных за движение и взаимодействие мышц. Сущ-ют исследования, связывающие с недостатком карнитина детский церебральный паралич и феномен "смерти в колыбели". 3 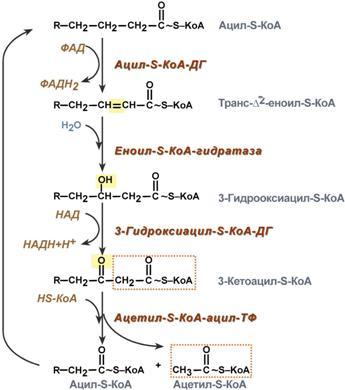 . После св-ия с карнитином ж к-та переносится ч/з мембрану транслоказой. Здесь на внутренней стороне мембраны фермент карнитин-ацилтрансфераза II вновь обр-ет ацил-S-КоА кот вступает на путь β-окисления. . После св-ия с карнитином ж к-та переносится ч/з мембрану транслоказой. Здесь на внутренней стороне мембраны фермент карнитин-ацилтрансфераза II вновь обр-ет ацил-S-КоА кот вступает на путь β-окисления.4. Процесс собственно β-окисления состоит из 4-х реакций, повторяющихся циклически. В них последовательно происходит окисление (ацил-SКоА-дегидрогеназа), гидратирование (еноил-SКоА-гидратаза) и вновь окисление 3-го атома углерода (гидроксиацил-SКоА-дегидрогеназа). В последней, трансферазной, реакции от жирной кислоты отщепляется ацетил-SКоА. К оставшейся (укороченной на два углерода) жирной кислоте присоединяется HS-КоА, и она возвращается к первой реакции. Все повторяется до тех пор, пока в последнем цикле не образуются два ацетил-SКоА. Расчет энергетического баланса β-окисления: При расчете количества АТФ, образуемого при β-окислении жирных кислот необходимо учитывать: *количество образуемого ацетил-SКоА – определяется обычным делением числа атомов углерода в жирной кислоте на 2; * 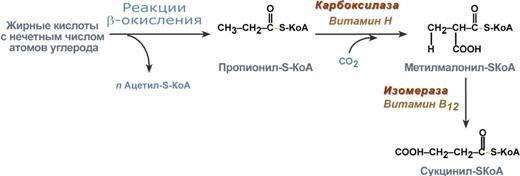 число циклов β-окисления. Число циклов β-окисления легко определить исходя из представления о жирной кислоте как о цепочке двухуглеродных звеньев. Число разрывов между звеньями соответствует числу циклов β-окисления. Эту же величину можно подсчитать по формуле (n/2 -1), где n – число атомов углерода в кислоте, число циклов β-окисления. Число циклов β-окисления легко определить исходя из представления о жирной кислоте как о цепочке двухуглеродных звеньев. Число разрывов между звеньями соответствует числу циклов β-окисления. Эту же величину можно подсчитать по формуле (n/2 -1), где n – число атомов углерода в кислоте, * 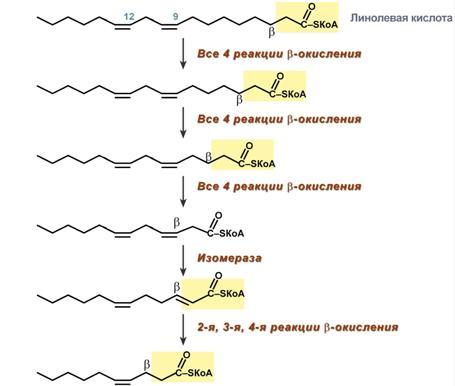 число двойных связей в жирной кислоте. В первой реакции β-окисления происходит образование двойной связи при участии ФАД. Если двойная связь в жирной кислоте уже имеется, то необходимость в этой реакции отпадает и ФАДН2 не образуется. Количество необразованных ФАДН2 соответствует числу двойных связей. Остальные реакции цикла идут без изменений; число двойных связей в жирной кислоте. В первой реакции β-окисления происходит образование двойной связи при участии ФАД. Если двойная связь в жирной кислоте уже имеется, то необходимость в этой реакции отпадает и ФАДН2 не образуется. Количество необразованных ФАДН2 соответствует числу двойных связей. Остальные реакции цикла идут без изменений; *количество энергии АТФ, потраченной на активацию (всегда соответствует двум макроэргическим связям). Окисление жирных кислот с нечетным числом углеродных атомов. Ж к-ты с нечетным числом углеродов поступают в организм с растительной пищей и морепродуктами. Их окисление происходит по обычному пути до последней реакции, в которой обр-ся пропионил-SКоА. Суть превращений пропионил-SКоА сводится к его карбоксилированию, изомеризации и обр-ию сукцинил-SКоА. В этих реакциях участвуют биотин и В12. Окисление ненасыщенных жирных кислот. При окислении ненасыщенных жирных кислот возникает потребность клетки в дополнительных ферментах изомеразах. Эти изомеразы перемещают двойные связи в жирнокислотных остатках из γ- в β-положение и переводят природные двойные связи из цис- в транс-положение. Т.о., уже имеющаяся двойная связь готовится к β-окислению и пропускается первая реакция цикла, в которой участвует ФАД. Глицерин – трехатомный спирт, водорастворим и легко всасывается из кишечника и по воротной вене поступает в печень. Окисление глицерина Глицерин сначала фосфорилируется с участием АТФ до глицерофосфата (3-фосфоглицерол). Затем под действием НАД-зависимой глицерофосфатдегидрогеназы окисляется до 3-фосфоглицеринового альдегида. Фосфоглицериновый альдегид далее может окисляться до пировиноградной и молочной кислоты. Связь в-окисления с ЦК и Дц: Таким образом, молекула жирной кислоты в конце концов распадается до продуктов, имеющих всего два углеродных атома, превращающихся в цикле трикарбоновых кислот. Восстановленные коферменты впоследствии вновь окисляются в дыхательной цепи с одновременным образованием макроэргических фосфатов. С точки зрения образования АТР, окисление жирных кислот составляет основной энергетический резерв организма. Регуляция в-окисления: Ключевой фермент – карнитинацилтрансфераза1, аллостерический фермент, в печени его аллостерический ингибитор – малонилКоА. Активируют: катехоламины, СТГ, глюкагон. Ингибирует: инсулин. Е значение в-окисление имеет для скелетных мышц (50% Е), для сердечной мышцы (70%), головной мозг и другие нервные ткани, а также эритроциты не используют жирные к-ты для окисления; они не поступают в головной мозг, т.к. не проходят ч/з гематоэнцефалический барьер. 23. Липолиз и липогенез. Значение. Зависимость липогенеза от ритма питания и состава пищи. Регуляция липолиза и липогенеза. Транспорт и использование жирных кислот, образующихся при мобилизации жира. Синтезируясь во время и сразу после приема пищи (липогенез) и запасаясь в жировой ткани, триацилглицеролы являются формой хранения насыщенных и мононенасыщенных жирных кислот. Распад триацилглицеролов (триглицеридов) по-другому называется липолиз или мобилизация жира. Он идет в жировых клетках постоянно и обычно существует равновесие между синтезом и распадом ТАГ. Даже в состоянии покоя организма печень, сердце, скелетные мышцы и другие ткани (кроме эритроцитов и нейроцитов) более 50% энергии получают из окисления жирных кислот, поступающих из жировой ткани благодаря фоновому липолизу. По мере уменьшения резервов глюкозы клетки все больше энергии получают из окисления жирных кислот. Таким образом, насыщенные жирные кислоты выполняют роль своеобразного буфера энергии в организме. Мобилизация триацилглицеролов и окисление жирных кислот активируется при нормальных физиологических стрессовых ситуациях – эмоциональный стресс, мышечная работа, голодание, при патологических состояниях – сахарный диабет I типа, другие гормональные заболевания (гиперкортицизм, гипертиреоз). В результате липолиза в адипоцитах образуются свободный глицерол и жирные кислоты. Глицерол с кровью доставляется в печень и почки, здесь фосфорилируется и окисляется в метаболит гликолиза диоксиацетонфосфат. В зависимости от условий ДАФ может включаться в реакции глюконеогенеза (при голодании, мышечной нагрузке) или окисляться в гликолизе до пировиноградной кислоты. Жирные кислоты транспортируются в крови в комплексе с альбуминами плазмы: -при физической нагрузке – в мышцы, -в обычных условиях и при голодании – в мышцы и большинство тканей, однако при этом около 30% жирных кислот захватывается печенью. При голодании и физической нагрузке после проникновения в клетки жирные кислоты вступают на путь β-окисления. В целом мобилизацию жира можно представить как последовательность следующих событий: 1.Липолиз – гормонзависимый распад ТАГ в жировой ткани или резервных ТАГ в самой клетке. 2.Транспорт жирных кислот из жировой ткани по крови в комплексе с альбумином. 3.Проникновение жирной кислоты в цитозоль клетки-мишени. 4.Активация жирной кислоты через присоединение HS-КоА. 5.Карнитин-зависимое перемещение жирной кислоты в митохондрию. 6.Окисление жирной кислоты с образованием ацетильных групп (в форме ацетил-S-КоА). 7.Сгорание ацетил-S-КоА в цикле лимонной кислоты или синтез (только в печени) кетоновых тел. В 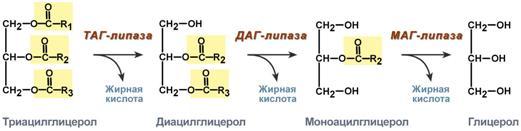 качестве источника энергии могут использоваться только свободные, т.е. неэстерифицированные, жирные кислоты. Поэтому триглицериды сначала гидролизуются при помощи специфических тканевых ферментов – липаз – до глицерина и свободных жирных кислот. Последние из жировых депо могут переходить в плазму крови (мобилизация высших жирных кислот), после чего они используются тканями и органами тела в качестве энергетического материала. В жировой ткани содержится несколько липаз, из которых наибольшее значение имеют триглицеридлипаза (так называемая гормоночувствитель-ная липаза), диглицеридлипаза и моноглицеридлипаза. Активность двух последних ферментов в 10–100 раз превышает активность первого. Три-глицеридлипаза активируется рядом гормонов (например, адреналином, норадреналином, глюкагоном и др.), тогда как диглицеридлипаза и мо-ноглицеридлипаза не чувствительны к их действию. Триглицеридлипаза является регуляторным ферментом. Первое, что происходит при использовании нейтрального жира во время голодания и физической нагрузки – это активация фермента, отвечающего за отщепление первой жирной кислоты от триацилглицерола. Фермент называется гормон-чувствительная триацилглицерол-липаза или ТАГ-липаза. Кроме ТАГ-липазы, в адипоцитах имеются еще диацилглицерол-липаза ( ДАГ-липаза) и моноацилглицерол-липаза (МАГ-липаза), активность которых высока и постоянна, однако в покое эта активность не проявляется из-за отсутствия субстрата. Как только в клетке после работы ТАГ-липазы появляются диацилглицеролы начинает работать постоянно активная ДАГ-липаза, продукт ее реакции моноацилглицерол (МАГ) является субстратом для МАГ-липазы. Образующиеся жирные кислоты и глицерол покидают клетку. Для регуляции активности ТАГ-липазы обязательно наличие гормонального влияния (адреналин, глюкагон, соматотропин, инсулин и др). качестве источника энергии могут использоваться только свободные, т.е. неэстерифицированные, жирные кислоты. Поэтому триглицериды сначала гидролизуются при помощи специфических тканевых ферментов – липаз – до глицерина и свободных жирных кислот. Последние из жировых депо могут переходить в плазму крови (мобилизация высших жирных кислот), после чего они используются тканями и органами тела в качестве энергетического материала. В жировой ткани содержится несколько липаз, из которых наибольшее значение имеют триглицеридлипаза (так называемая гормоночувствитель-ная липаза), диглицеридлипаза и моноглицеридлипаза. Активность двух последних ферментов в 10–100 раз превышает активность первого. Три-глицеридлипаза активируется рядом гормонов (например, адреналином, норадреналином, глюкагоном и др.), тогда как диглицеридлипаза и мо-ноглицеридлипаза не чувствительны к их действию. Триглицеридлипаза является регуляторным ферментом. Первое, что происходит при использовании нейтрального жира во время голодания и физической нагрузки – это активация фермента, отвечающего за отщепление первой жирной кислоты от триацилглицерола. Фермент называется гормон-чувствительная триацилглицерол-липаза или ТАГ-липаза. Кроме ТАГ-липазы, в адипоцитах имеются еще диацилглицерол-липаза ( ДАГ-липаза) и моноацилглицерол-липаза (МАГ-липаза), активность которых высока и постоянна, однако в покое эта активность не проявляется из-за отсутствия субстрата. Как только в клетке после работы ТАГ-липазы появляются диацилглицеролы начинает работать постоянно активная ДАГ-липаза, продукт ее реакции моноацилглицерол (МАГ) является субстратом для МАГ-липазы. Образующиеся жирные кислоты и глицерол покидают клетку. Для регуляции активности ТАГ-липазы обязательно наличие гормонального влияния (адреналин, глюкагон, соматотропин, инсулин и др).В целом последовательность событий активации липолиза выглядит следующим образом: 1.Молекула гормона (адреналин, глюкагон, АКТГ) взаимодействует со своим рецептором. 2.Активный гормон-рецепторный комплекс воздействует на мембранный G-белок. 3.G-белок активирует фермент аденилатциклазу. 4.Аденилатциклаза превращает АТФ в цАМФ – вторичный посредник (мессенджер). 5. цАМФ аллостерически активирует фермент протеинкиназу А. 6.Протеинкиназа А фосфорилирует ТАГ-липазу и активирует ее. 7.ТАГ-липаза отщепляет от триацилглицеролов жирную кислоту в 1 или 3 положении с образованием диацилглицерола (ДАГ).А 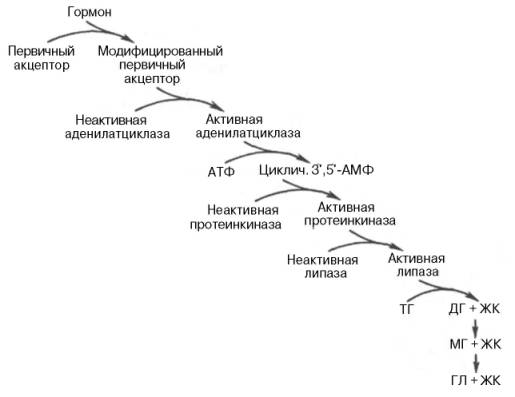 ктивная триглицеридлипаза расщепляет триглицерид на диглицерид и жирную кислоту. Затем при действии ди- и моноглицеридлипаз образуются конечные продукты липо-лиза – глицерин и свободные жирные кислоты, которые поступают в кровяное русло. ктивная триглицеридлипаза расщепляет триглицерид на диглицерид и жирную кислоту. Затем при действии ди- и моноглицеридлипаз образуются конечные продукты липо-лиза – глицерин и свободные жирные кислоты, которые поступают в кровяное русло.Скорость липолиза триглицеридов не является постоянной, она подвержена регулирующему влиянию различных факторов, среди которых особое значение имеют нейрогормональные. Связанные с альбуминами плазмы крови в виде комплекса свободные жирные кислоты с током крови попадают в органы и ткани, где комплекс распадается, а жирные кислоты подвергаются либо β-окислению, либо частично используются для синтеза триглицеридов, глицерофосфолипидов, сфингофосфолипидов и других соединений, а также на эстерификацию холестерина. При физической работе и других состояниях организма, требующих повышенных энергозатрат, потребление триглицеридов жировой ткани как энергетического резерва увеличивается. Активируют: катехоламины (адреналин), глюкагон, тироксин, кортизол, цАМФ. Ингибирует: инсулин. Л 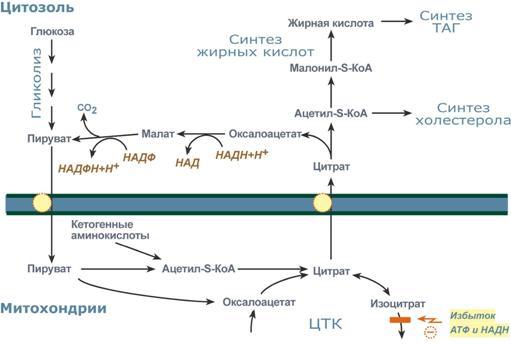 ипогенез – это синтез жира из глицерина и жирных к-т. Происходит в печени и жировой ткани. Глицерин и жирные к-ты должны быть в активной форме. В жировой тк глицеролкиназа отсутствует, поэтому активная форма обр-ся из ДОАФ – метаболита гликолиза (из глюкозы). Синтез жира протекает в адбсортивный период (после еды). Молекулы жира в адипоцитах объединяются крупной каплей и явл-ся компактной формой хранения Е запасов. Обмен жира в жировой тк протекает очень активно: обновляется полностью за несколько дней. В печени ежесуточно обр-ся 20-50г жира, который поступает в кровь в составе ЛПОНП. Ключевой фермент: ацилтрансфераза. Активируют: инсулин, эстрогены и АТФ. Ингибируют: катехоламины, СТГ, йодтиронины, АКТГ, АДФ. Синтез нейтральных жиров происходит за счет этерификации глицеро-1-фосфата двумя активированными жирными кислотами. Фосфатная группа образованной фосфатидной кислоты отщепляется фосфатазами, что приводит к образованию диацилглицерина, который далее реагирует с другой активированной жирной кислотой, образуя триацилглицерин. Триацилглицерины составляют энергетическое депо организма. Они обладают очень высокой теплотой окисления, равной 37,6 кДж/моль. Жиры локализованы в жировых клетках (адипоцитах) и характеризуются высокой скоростью метаболизма. Их превращение непосредственно регулируется гормонами, в частности инсулином и адреналином. Реакции биосинтеза липидов могут идти в гладкой эндоплазматической сети клеток всех органов. Субстратом для синтеза жиров de novo является глюкоза. Как известно, попадая в кл, глюкоза превращается в гликоген, пентозы и окисляется до пировиноградной кислоты. При высоком поступлении глюкоза используется для синтеза гликогена, но этот вариант ограничивается объемом клетки. Поэтому глюкоза "проваливается" в гликолиз и превращается в пируват либо напрямую, либо через пентозофосфатный шунт. Во втором случае образуется НАДФН, который понадобится впоследствии для синтеза жирных к-т. Пируват переходит в митохондрии и декарбоксилируется в ацетил-SКоА и вступает в ЦТК. Однако в состоянии покоя, при отдыхе, при наличии избыточного количества энергии в клетке реакции ЦТК (в частности, изоцитратдегидрогеназная реакция) блокируются избытком АТФ и НАДН. В результате накапливается первый метаболит ЦТК – цитрат. По градиенту концентрации он перемещается в цитозоль, расщепляется с образованием ацетил-SКоА, который далее используется в биосинтезе холестерола, жирных кислот и триацилглицеролов. Оксалоацетат, также образуемый из цитрата, восстанавливается до яблочной кислоты и возвращается в митохондрии *посредством малат-аспартатного челнока (на рисунке не показан), *после декарбоксилирования малата до пирувата НАДФ-зависимым малик-ферментом. Образованный НАДФН будет использован при синтезе жирных кислот. ипогенез – это синтез жира из глицерина и жирных к-т. Происходит в печени и жировой ткани. Глицерин и жирные к-ты должны быть в активной форме. В жировой тк глицеролкиназа отсутствует, поэтому активная форма обр-ся из ДОАФ – метаболита гликолиза (из глюкозы). Синтез жира протекает в адбсортивный период (после еды). Молекулы жира в адипоцитах объединяются крупной каплей и явл-ся компактной формой хранения Е запасов. Обмен жира в жировой тк протекает очень активно: обновляется полностью за несколько дней. В печени ежесуточно обр-ся 20-50г жира, который поступает в кровь в составе ЛПОНП. Ключевой фермент: ацилтрансфераза. Активируют: инсулин, эстрогены и АТФ. Ингибируют: катехоламины, СТГ, йодтиронины, АКТГ, АДФ. Синтез нейтральных жиров происходит за счет этерификации глицеро-1-фосфата двумя активированными жирными кислотами. Фосфатная группа образованной фосфатидной кислоты отщепляется фосфатазами, что приводит к образованию диацилглицерина, который далее реагирует с другой активированной жирной кислотой, образуя триацилглицерин. Триацилглицерины составляют энергетическое депо организма. Они обладают очень высокой теплотой окисления, равной 37,6 кДж/моль. Жиры локализованы в жировых клетках (адипоцитах) и характеризуются высокой скоростью метаболизма. Их превращение непосредственно регулируется гормонами, в частности инсулином и адреналином. Реакции биосинтеза липидов могут идти в гладкой эндоплазматической сети клеток всех органов. Субстратом для синтеза жиров de novo является глюкоза. Как известно, попадая в кл, глюкоза превращается в гликоген, пентозы и окисляется до пировиноградной кислоты. При высоком поступлении глюкоза используется для синтеза гликогена, но этот вариант ограничивается объемом клетки. Поэтому глюкоза "проваливается" в гликолиз и превращается в пируват либо напрямую, либо через пентозофосфатный шунт. Во втором случае образуется НАДФН, который понадобится впоследствии для синтеза жирных к-т. Пируват переходит в митохондрии и декарбоксилируется в ацетил-SКоА и вступает в ЦТК. Однако в состоянии покоя, при отдыхе, при наличии избыточного количества энергии в клетке реакции ЦТК (в частности, изоцитратдегидрогеназная реакция) блокируются избытком АТФ и НАДН. В результате накапливается первый метаболит ЦТК – цитрат. По градиенту концентрации он перемещается в цитозоль, расщепляется с образованием ацетил-SКоА, который далее используется в биосинтезе холестерола, жирных кислот и триацилглицеролов. Оксалоацетат, также образуемый из цитрата, восстанавливается до яблочной кислоты и возвращается в митохондрии *посредством малат-аспартатного челнока (на рисунке не показан), *после декарбоксилирования малата до пирувата НАДФ-зависимым малик-ферментом. Образованный НАДФН будет использован при синтезе жирных кислот. |