Гилберт С. Биология развития. Т.1.doc , БИР. Библиография Гилберт С. Биология развития в 3х т. Т. I Пер с англ. М. Мир, 1993. 228 с
 Скачать 10.9 Mb. Скачать 10.9 Mb.
|

Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.______________ ОПЛОДОТВОРЕНИЕ: ВОЗНИКНОВЕНИЕ НОВОГО ОРГАНИЗМА 45
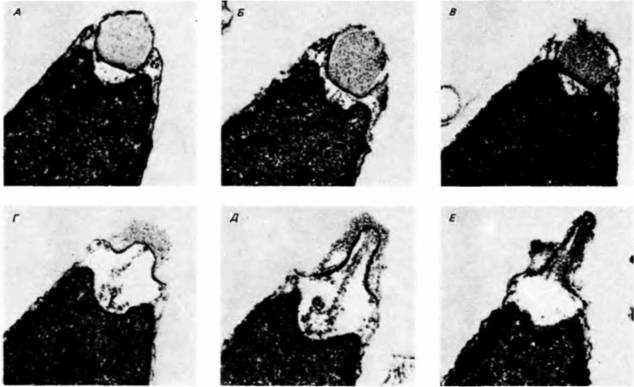
в образовании акросомного выроста (рис. 2.12). Этот вырост возникает в результате полимеризации молекул глобулярного актина и образования актиновых филаментов процесс, вызываемый, как полагают, выходом ионов водорода из головки спермия (Tilney et al., 1978; Schackmann et al., 1978). До активации спермия полимеризация актина, вероятно, блокирована регуляторным белком: сопровождающее активацию повышение внутриклеточного pH может воспрепятствовать выполнению этой функции. Значительное повышение pH вызывает не только образование акросомного выроста, но и активирует динеиновую АТФазу в шейке спермия. В результате такой активации происходит быстрое использование АТФ и 50%-ное увеличение интенсивности дыхания митохондрий. Произведенная энергия используется главным образом для обеспечения движений жгутика (Tombes, Shapiro, 1985). Действующее начало студенистой оболочки, инициирующее у морских ежей акросомную реакцию, часто бывает высокоспецифичным. Спермии морских ежей ArbaciapunctulataиStrongylocentrotusdroebachiensisреагируют только на воздействие материала студенистой оболочки яиц собственного вида. Однако спермии S. purpuratusмогут быть активированы материалом студенистой оболочки яиц не только собственного вида, но и Lytechinusvariegalux(но не A. punctulata) (Summers, Hylander, 1975). Следовательно, студенистая оболочка у некоторых видов (но не у всех) может обеспечивать видоспецифическое узнавание гамет. Узнавание спермия и яйца: контакт между гаметамиВидоспецифическое узнавание у морских ежейСразу же после того, как спермий морского ежа проник в студенистую оболочку яйца, его акросомный вырост вступает в контакт с наружной поверхностью желточной оболочки (рис. 2.13). Основной этап видоспецифического узнавания осуществляется в этот момент. Белок акросомы, при посредстве которого происходит это узнавание, называется биндином. В 1977 г. Вакье с сотрудниками (Vacquier, Moy, 1977) изолировали этот нерастворимый белок с молекулярной массой 30 500 дальтон из акросомы спермиев Strongylocentrotuspurpuratus: биндин способен Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.46 ГЛАВА 2
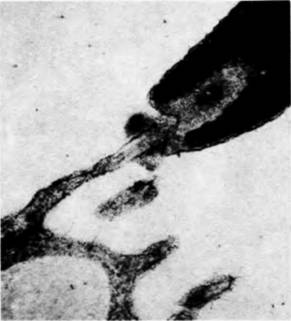
прикрепляться к яйцам S. purpuratus, освобожденным от студенистой оболочки, и к изолированным желточным оболочкам (рис. 2.14). Взаимодействие биндина с яйцом видоспецифично (Glabe, Vacquier. 1977; Glabe, Lennarz, 1979); этот белок, выделенный из акросомы спермиев S. purpuratus, вызывает агглютинацию яиц своею вида, освобожденных от студенистой оболочки, но не яиц близкородственного вида S. franciscanus. Было обнаружено (Моу, Vacquier, 1979), что локализация биндина строго специфична он находится на акросомном выросте, именно там, где он должен находиться, чтобы обеспечить узнавание спермием и яйцом друг друга. Использованный этими авторами метод иммунохимического окрашивания биндина представлен на рис. 2.15. А. Антитела к очищенному биндину морского ежа получали, инъецируя биндин кроликам. Спермии морского ежа, осуществившие акросомную реакцию, связывали этот антибиндин. После отмывания непрореагировавших антител спермии обрабатывали антителами свиньи, ковалентно связанными с пероксидазами и способными присоединяться к антителам кролика. В результате молекулы пероксидазы оказывались везде, где присутствовал биндин. Пероксидаза катализирует образование темного осадка при взаимодействии диаминобензидина и перекиси водорода. Когда к обработанным спермиям добавляли эти два субстрата, осадок покрывал акросомный вырост (рис. 2.15, Б). Более того, до осуществления акросомной реакции выявить биндин на поверхности спермия не удавалось, а затем его обнаруживали в месте соединения спермия с яйцом (рис. 2.15. В). Данные биохимических исследований показали, что биндины близкородственных видов морских ежей на самом деле различны. Эти данные заставляют предположить существование на желточной оболочке видоспецифических рецепторов для связывания биндина. О том же свидетельствуют опыты по насыщению яиц морского ежа спермиями (Vacquier. Payne, 1973). Как видно на рис. 2.16. спермии прикрепляются к яйцу не на всей его поверхности. Даже в случаях насыщения спермиями (приблизительно 1500 спермиев на 1 яйцо) на поверхности желточной оболочки остаются свободные участки, что говорит об ограниченном числе мест для прикрепления спермиев. Из желточных оболочек яиц морского ежа был выделен крупный гликопротеиновый комплекс, который видоспецифическим образом связывал радиоактивный биндин (Glabe, Vacquier, 1978; Rossignol et al., 1984). Этот гликопротеин способен конкурировать с яйцами за спермии того же вида. Таким образом, если спермии S. purpuratus смешать с биндиновыми рецепторами из желточных оболочек S. purpuratus, то спермии прикрепятся к ним и не будут оплодотворять яйца. Однако выделенные биндиновые рецепторы S. purpuraïus не препятствуют оплодотворению яиц родственных видов морских ежей. Следовательно, видоспецифическое узнавание гамет морского ежа происходит на двух уровнях — на уровне активации акросомной реакции и на уровне прикрепления спермия к желточной оболочке. Соединение гамет у млекопитающихДо сих пор наше обсуждение касалось тех организмов, у которых оплодотворение происходит во внешней среде. У млекопитающих оплодотворение внутреннее и процесс оплодотворения приспособлен к внутренней среде организма. И в самом деле, было обнаружено, что у млекопитающих половые пути самки принимают активное участие в процессе оплодотворения. Спермии млекопитающих сразу после эякуляции не способны к акросомной реакции; для того чтобы эта реакция стала возможной, спермиям необходимо в течение некоторого времени находиться в половых путях самки. Условия, требующиеся для капацитации (приобретения спермиями оплодотворяющей способности), варьируют в зависимости от вида (Gwatkin, 1976); пребывание спермиев в половых путях самки может быть заменено их инкубацией in vitro в жидком содержимом яйцеводов или матки. Одна из гипотез относительно природы процесса капацитации постулирует, что процесс этот заключается в изменениях структуры липидов клеточ- |