Гилберт С. Биология развития. Т.1.doc , БИР. Библиография Гилберт С. Биология развития в 3х т. Т. I Пер с англ. М. Мир, 1993. 228 с
 Скачать 10.9 Mb. Скачать 10.9 Mb.
|

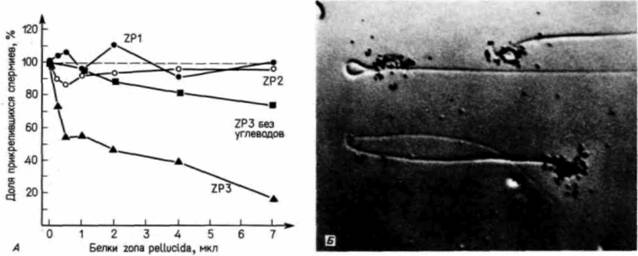
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с._______________ ОПЛОДОТВОРЕНИЕ: ВОЗНИКНОВЕНИЕ НОВОГО ОРГАНИЗМА _____________ 49 ми этой оболочки. В ном опыте активным конкурентом яйца является гликопротеин с молекулярной массой 83000 дальтон (ZP3), выделенный из прозрачной оболочки (Bleil, Wassarman. 1980, 1986; см. рис. 2.17). Таким образом, в прозрачной оболочке содержится особый гликопротеин, к которому прикрепляется спермий. После того как прикрепление произошло, тот же самый белок, как было показано, возбуждает акросомную реакцию. Это позволяет спермию мыши концентрировать свои протеолитические ферменты непосредственно в месте прикрепления к прозрачной оболочке. Спермии всех млекопитающих содержат лизин, разрушающий прозрачную оболочку в месте прикрепления (Stambaugh, Buckley. I960). Этот лизин, называемый акрозином, по аминокислотной последовательности очень сходен с трипсином, и чтобы начать работать, он, подобно трипсину, должен быть активирован. Было установлено (Wincek et al., 1979; Stambaugh, Mastroianni, 1980), что акрозин, находящийся в акросомном пузырьке, неактивен, но под воздействием гликопротеина из половых путей самки происходит его активация. Здесь мы опять убеждаемся в том, что половые пути самки являются не просто каналами для проведения спермы, но и активными участниками процесса оплодотворения. Молекулярный механизм, с помощью которого прозрачная оболочка яйца и спермий млекопитающих узнают друг друга, полностью еще не изучен. Распространенная гипотеза, предложенная для объяснения механизма соединения гамет у млекопитающих, постулирует существование на поверхности спермия белка, способного узнавать видоспецифический углевод гликопротеина прозрачной оболочки (Wassarman, 1987). Показано (Horrnun et al., 1984; Florman. Wassarman, 1985), что у мыши важную роль в узнавании спермия прозрачной оболочкой Играет углеводная группа на гликопротеине ZP3. После удаления этих углеводных групп ZP3 утрачивает способность конкурировать за прикрепление спермия (однако для инициации акросомной реакции, по-видимому, необходима вся молекула гликопротеина). В лаборатории Шура (Shur, Hall, 1982a.b; Lopez et al., 1985) с использованием другого подхода было показано, что рецепторы спермия для прозрачной оболочки представляют собой фермент на поверхности его головки, который узнает сахар N-ацетилглюкозамин (рис. 2.18.4 ). Этот фермент (N-ацетилглюкозамин:галактозилтрансфераза) встроен в плазматическую мембрану спермия непосредственно над акросомой, причем его активный центр обращен наружу. Эта гликозилтрансфераза, вероятно, катализирует присоединение галактозы к углеводной цепи, оканчивающейся N-ацетилглюкозамином. Однако в половых путях самки отсутствуют активированные остатки галактозы. Поэтому, хотя фермент может присоединяться к N-ацетилглюкозаминовым остаткам белков zona pellucida (возможно, к ZP3. что, однако, еще не установлено) точно так же, как любой фермент соединяется с субстратом, он не может катализировать реакцию, поскольку второй ее участник отсутствует. В результате ферменты (на спермии) остаются связанными со своими субстратами (на яйце).
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.50________________ ГЛАВА 2______________________________________________________________________________
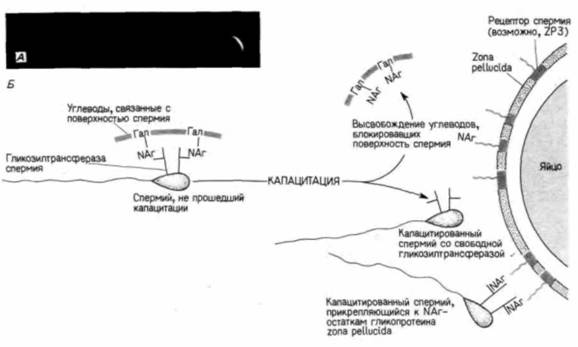
Если приведенная гипотеза справедлива, то прикрепление спермия можно предотвратить ингибированием фермента или добавлением второго участника реакции – активированной галактозы. Именно такой результат и был получен Шуром с сотрудниками: как при ингибировании гликозилтрансферазы (антителами), так и при завершении реакции (путем добавления УДФ-галактозы) спермий к яйцу не прикреплялся. Эти данные позволили связать обнажение гликозилтрансферазы на поверхности спермия с его капацитацией. Уже около десятка лет известно, что одно из проявлений капацитации — приобретение спермием способности узнавать прозрачную оболочку яйца. На поверхности спермия, не прошедшего капацитации, находятся углеводы ("coating factors"), которые препятствуют его прикреплению к яйцу. При капацитации эти углеводы удаляются, а когда их добавляют к капацитированным спермиям, то спермии утрачивают способность прикрепляться к яйцам. Углеводы, блокирующие прикрепление спермия, представляют собой, по-видимому, полимер из повторяющихся остатков галактозы и N-ацетилтлюкозамина — именно тот тип молекулы, который способен связывать активный центр гликозилтрансферазы в головке спермия. Объединение накопленных данных позволяет создать модель, представленную на рис. 2.18.Б. Прозрачная оболочка выступает как носитель субстрата остатков N-ацетилглюкозамина, а спермий - молекул гликозилтрансферазы, которые к этим остаткам прикрепляются. Капацитация обнажает гликозилтрансферазы на поверхности спермия, и их соединение с N-ацетилглюкозаминовыми остатками прозрачной оболочки по типу ключ-замок прикрепляет спермий к яйцу. Такая же гликозилтрансфераза была обнаружена в спермиях морской свинки и человека, но ее роль в соединении гамет у этих видов предстоит еще выяснить. Слияние гамет и предотвращение полиспермииСлияние плазматических мембран спермия и ооцитаВслед за узнаванием спермия желточной (или прозрачной) оболочкой яйца происходит лизис |