|
Гилберт С. Биология развития. Т.1.doc , БИР. Библиография Гилберт С. Биология развития в 3х т. Т. I Пер с англ. М. Мир, 1993. 228 с
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.
|
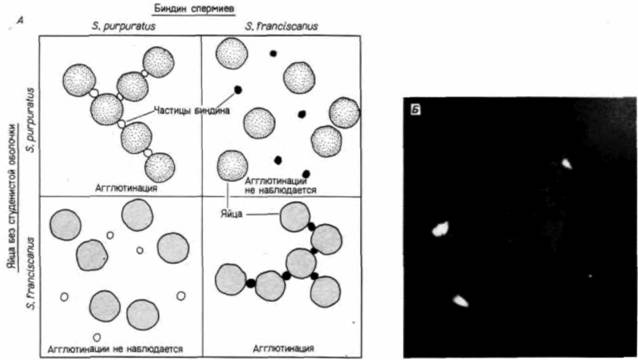
Рис. 2.14. Видоспецифическая агглютинация яиц, освобожденных от студенистой оболочки, при добавлении биндина А, Агглютинацию вызывает добавление 213 mkг биндина в пластмассовую чашку, содержащую 0.25 мл 2% -ной (по объему) суспензии яиц. В течение 2-5 мин чашку осторожно покачивают и затем фотографируют (пояснительная схема сделана по фотографиям Glabe, Vacquier, 1977). Б. Микрофотография флуоресцирующих яиц S. purpuratus, соединенных друг с другом частицами биндина S. purpuratus, меченными флуоресцеином. В местах, где два яйца соприкасались, неизменно присутствовали частицы биндина. (Из Glabe, Lennarz, 1979: фотография с любезного разрешения авторов.)
|
|
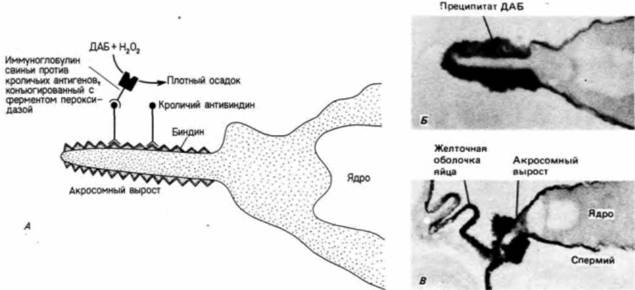
Рис. 2. 15. Локализация биндина на акросомном выросте. А, Иммунохимический метод выявления локализации биндина. Кроличьи антитела взаимодействуют с обнажившимся биндином. Затем к этим антителам добавляют ковалентно связанные с молекулой пероксидазы антитела свиньи против кроличьих антител. Пероксидаза катализирует реакцию между диаминобензидином (ДАВ) и перекисью, в результате которой образуется электроноплотный осадок. Таким образом, этот осадок будет возникать только там, где имеется биндин. Б.Локализация биндина на акросомном выросте после акросомной реакции (х 133 200). В. Локализация биндина на акросомном выросте в месте соединения спермия с яйцом. (Б и В -- из Moy,. Vacquier. 1979; фотографии с любезного разрешения V. D. Vacquier.)
|
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.
48 ГЛАВА 2
|
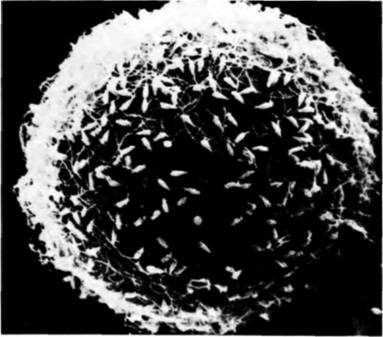
Рис. 2.16. Микрофотография спермиев морского ежа, прикрепившихся к желточной оболочке яйца, полученная с помощью сканирующего электронного микроскопа. (Фотография с любезного разрешения С. Glabe, L. Perez. W.J. Lennarz.)
|
ной мембраны спермии. Это представление основывается на исследованиях (Davis et al., 1980), которые показывают, что соотношение холестерин : фосфолипиды в мембране спермия по мере капацитации снижается и что молекулы альбумина, имеющиеся в половых путях самки, способны отнимать холестерин у спермия. Продолжительность капацитации различна у разных видов и тесно коррелирует с временем, требующимся для изменения соотношения холестерин:фосфолипиды (Davis. 1981). Полагают, что снижение содержания холестерина дестабилизирует мембрану акросомного пузырька и что без таких изменений липидного состава слияние мембран, наблюдающееся при акросомной реакции, не может осуществиться. Вместе с тем при капацитации с поверхности спермия удаляются особые, связанные с ней вещества ("coating factors"), которые, оставаясь на поверхности, препятствуют оплодотворению (Yanagimachi. 1981).
Вопрос о том, как инициируется акросомная реакция у млекопитающих, остается спорным: местонахождение спермиев в момент ее инициации у разных видов может быть различным. Это различие, вероятно, сопряжено с разной способностью спермиев проникать через лучистый венец, окружающий яйцо. У некоторых видов акросомная реакция происходит, по-видимому, в то время, когда спермий находится на некотором расстоянии от яйца. Так, например, по имеющимся данным, у кролика акросомная реакция может быть индуцирована растворимыми веществами, выделяемыми яйцом (Bedford, 1968), тогда как у морской свинки она происходит в определенное время независимо от присутствия яиц (Yanagimachi. Usui, 1974). В этих случаях акросомная реакция приводит к выделению ферментов, лизирующих связи между клетками лучистого венца (Zaneveld, Williams. 1970; Williams. 1972). Выделенные ферменты, называемые лизинами, включают гиалуронидазу, расщепляющую гиалуроновую кислоту, которая составляет большую часть внеклеточного матрикса. и фермент, рассеивающий клетки лучистого венца, специфическим образом разрушая связи между ними. С помощью этих ферментов спермий проходит через лучистый венец и достигает прозрачной оболочки яйца.
У наиболее хорошо изученных млекопитающих мыши и хомячка акросомная реакция происходит после прикрепления спермия к прозрачной оболочке (Saling et al., 1979; Florman, Storey. 1982; Cherr et al.. 1986). У этих животных спермии способны, по-видимому, проникать через лучистый венец, не выделяя больших количеств гиалуронидазы (Talbot at al.. 1985).
Прозрачная оболочка яиц млекопитающих выполняет такую же роль, как и желточная оболочка у иглокожих. Прикрепление спермия к прозрачной оболочке лишь относительно, а не абсолютно видоспецифично (видовая специфичность при внутреннем оплодотворении — не столь важная проблема). Спермии мыши можно лишить способности прикрепляться к прозрачной оболочке яиц своего вида, если их предварительно инкубировать с гликопротеина-
|
|
|
 Скачать 10.9 Mb.
Скачать 10.9 Mb.