Гилберт С. Биология развития. Т.1.doc , БИР. Библиография Гилберт С. Биология развития в 3х т. Т. I Пер с англ. М. Мир, 1993. 228 с
 Скачать 10.9 Mb. Скачать 10.9 Mb.
|

Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с._______________ ОПЛОДОТВОРЕНИЕ ВОЗНИКНОВЕНИЕ НОВОГО ОРГАНИЗМА 53
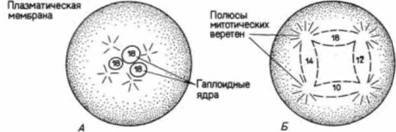
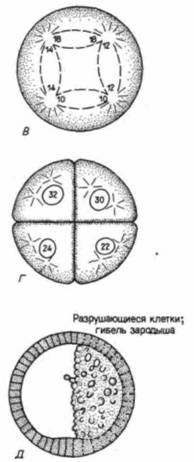
иллюстрирующей особую роль липидных молекул в слиянии плазматических мембран гамет. Слияние представляет собой активный процесс, часто опосредованный специфическими «фузогенными» (fusogenic) белками. Известно, что такие белки, как НА-белок вируса гриппа и F-белок вируса Сендай способствуют слиянию клеток: не исключено, что биндин относится к белкам именно такого типа. Сравнительно недавно было обнаружено (Glabe, 1985), что биндин морского ежа вызывает слияние фосфолипидных пузырьков между собой и что, подобно вирусным фузогенным белкам, биндин содержит около своего N-конца протяженный участок гидрофобных аминокислот. У брюхоногого моллюска Наliotisлизин, растворяющий желточную оболочку, также обладает фузогенной активностью (Hong. Vacquicr. 1986), а в головке спермиев морской свинки содержится белок, играющий важную роль в слиянии спермия и яйца у этого вида (Primakoff et al.. 1987). Таким образом, один из белков головки спермия, по-видимому, стимулирует слияние плазматических мембран спермия и яйца. Предотвращение полиспермииСпособность плазматической мембраны яйца к слиянию необходима для его соединения со спермием, однако, как только это слияние произойдет, такая способность становится опасным свойством. У морских ежей, как и у большинства изученных животных, каждый проникающий в яйцо спермий снабжает его гаплоидным ядром и центриолью. При нормальном моноспермном оплодотворении, когда в яйцо проникает только один спермий, гаплоидные ядра спермия и яйца соединяются, образуя диплоидное ядро оплодотворенною яйца (зиготы): при этом восстанавливается присущее данному виду число хромосом. Внесенная спермием центриоль делится и образует два полюса митотического веретена дробления. Проникновение множества спермиев — полиспермия у большинства животных приводит к гибельным последствиям. У морских ежей при оплодотворении двумя спермиями возникает триплоидное ядро, в котором каждая хромосома представлена не двумя, а тремя копиями. Такое оплодотворение приводит к вредным последствиям в особенности потому, что вместо нормального процесса — разделения хромосом с помощью биполярного веретена между двумя дочерними клетками — происходит распределение триплоидного набора между четырьмя клетками. А поскольку не существует механизма, который обеспечил бы получение каждой из четырех дочерних клеток нормального числа и всех необходимых типов хромосом, последние распределяются неравномерно: одни клетки получают лишние копии некоторых хромосом, тогда как у других эти хромосомы отсутствуют (рис. 2.21). Теодор Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.54 ГЛАВА 2 Бовери в 1902 г. показал, что такие клетки либо погибают, либо развиваются атипично. В процессе эволюции животные выработали разные способы, позволяющие избежать объединения более чем двух гаплоидных ядер. Самый обычным способ заключается в том, чтобы предотвратить проникновение в яйцо более чем одного спермия. В яйце морского ежа имеются два механизма зашиты от полиспермии: быстрая реакция, которая осуществляется путем электрических изменений в плазматической мембране, и более медленная реакция, вызванная экзоцитозом кортикальных гранул. БЫСТРЫЙ БЛОК ПОЛИСПЕРМИИ. Для оплодотворения необходимо, чтобы плазматическая мембрана яйца обладала способностью сливаться с мембраной спермия, однако она должна утрачивать эту способность сразу же после того, как первый спермий вступит с ней в контакт (Just. I9I9). Быстрый блок полиспермии достигает этой цели путем изменения электрического потенциала плазматической мембраны яйца. Клеточная мембрана представляет собой своего рода барьер между цитоплазмой яйца и внешней средой, обладающий свойством избирательности, и концентрация ионов по обе стороны плазматической мембраны сильно различается. Это различие особенно значительно для ионов натрия (Na+) и калия (К+). В морской воде концентрация ионов натрия очень велика, тогда как в цитоплазме яйца свободных ионов натрия почти нет. Для ионов калия соотношение обратное. Такое состояние поддерживается плазматической мембраной, которая устойчиво препятствует проникновению натрия в ооцит и утечке ионов калия в окружающую среду. Если один электрод ввести в яйцо, а второй электрод поместить вне его, то можно измерить постоянную разность потенциалов по обе стороны плазматической мембраны. Этот мембранный потенциал покоя, как правило, имеет величину, равную примерно 70 милливольт, что обычно выражают как -70 мВ, поскольку содержимое клетки отрицательно заряжено по отношению к внешней среде. В течение одной десятой секунды после при крепления первого спермия мембрана деполяризуется и мембранный потенциал достигает положительного уровня. Это результат поступления в яйцо небольшою количества ионов натрия, которое доводит разность потенциалов до 0 или даже + 20 мВ (рис. 2.22). Спермии способны сливаться с мембранами, обладающими потенциалом покоя —70 мВ, но неспособны сливаться с мембранами, потенциал которых имеет менее отрицательное значение, чем - 10 мВ. Неизвестно, каким образом прикрепление или проникновение спермия дает сигнал к открыванию натриевых каналов, однако недавно были получены данные (Gould. Stephano. 1987), которые
могут дать ключ к пониманию этого процесса: из спермиев Urechis был выделен акросомный белок, способный открывать натриевые каналы в неоплодотворенном яйце Urechis. Более того, когда яйца подвергали воздействию этого белка, скорость поступления натрия и возникавшее в результате его поступления смешение мембранного потенциала были сходны с аналогичными изменениями, вызываемыми живыми спермиями. Открывание натриевых каналов в яйце индуцируется, по-видимому, прикреплением к нему спермия. При изучении оплодотворения у морского ежа Lytechinus с точной регистрацией времени наступления разных процессов было установлено, что натрий поступает в яйцо до слияния плазматических мембран гамет (Longo et al.. 1986; Hinkley et al., 1986). Джеф (Jaffe, 1976) и ее сотрудники обнаружили, что можно индуцировать полиспермию путем искусственной фиксации потенциала на уровне -70 мВ и можно, наоборот, предотвратить оплодотворение, искусственно поддерживая потенциал на положительном уровне. Быстрый блок полиспермии можно также предотвратить, снижая концентрацию ионов натрия в воде (рис. 2.23). Если содержание ионов натрия недостаточно для того, чтобы вызвать смешение мембранною потенциала до положительных значений, то наблюдается полиспермия (Gould-Somero el al.. 1979; Jaffe, 1980). Электрический блок полиспермии существует также у лягушки (Cross, Elinson. 1980), но его, по-видимому, нет у млекопитающих (Jaffe, Cross. 1983). медленный блок полиспермии. Яйца морских ежей (и многих других животных) обладают еще одним механизмом для обеспечения моноспермности оплодотворения. Быстрый блок полиспермии действует недолго - мембранный потенциал в яйце морского ежа остается положительным только около одной минуты. Это кратковременное смещение потенциала может быть недостаточным для предотвращения полиспермии: было показано (Carroll. Epel. 1975). что если спермии, прикрепившиеся к жел- |