Гилберт С. Биология развития. Т.1.doc , БИР. Библиография Гилберт С. Биология развития в 3х т. Т. I Пер с англ. М. Мир, 1993. 228 с
 Скачать 10.9 Mb. Скачать 10.9 Mb.
|

Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.132 Г ЛАВА 4 ______________________
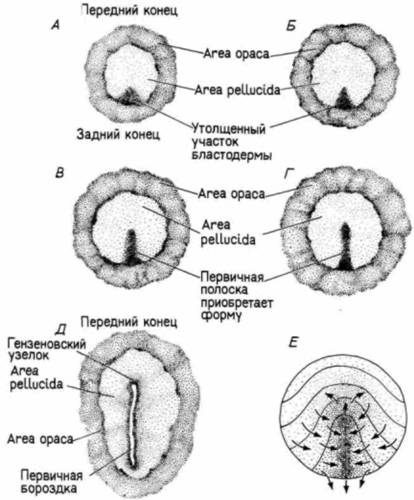
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с._______________ ГАСТРУЛЯЦИЯ: ПЕРЕРАСПРЕДЕЛЕНИЕ КЛЕТОК__________________________________________ 133 родыша птиц вступают в первичную полоску индивидуально. Вместо того чтобы соединиться в пласт тесно связанных между собой клеток, мигрирующие клетки образуют в бластоцеле рыхлое скопление мезенхимы Кроме того, в гаструле птиц не возникает настоящего архентерона. По мере поступления клеток в первичную полоску она удлиняется в направлении будущего головного конца зародыша. В то же самое время клетки вторичного гипобласта продолжают мигрировать вперед от заднего края бластодермы. Удлинение первичной полоски, по-видимому, коррелировано с этим движением клеток вторичного гипобласта. Первыми через первичную полоску мигрируют клетки, предназначенные стать передней кишкой. Эта ситуация вновь оказывается сходной с той, что наблюдалась у амфибий. Попав в бластоцель, эти клетки движутся вперед и в конце концов замещают собой клетки гипобласта в переднем отделе зародыша. Следующие клетки, вступающие в бластоцель через гензеновский узелок, также движутся вперед, но не распространяются в вентральном направлении в такой степени, как презумптивные энтодермальные клетки. Они остаются между энтодермой и эпибластом, становясь впоследствии клетками головной мезодермы и хордомезодермы (хорды). Эти рано выселяющиеся в бластоцель клетки движутся вперед, приподнимая над собой передний участок эпибласта по среднеспинной линии и образуя головной отросток. Между тем продолжается миграция клеток через первичную полоску. Попав в бластоцель. они подразделяются на два потока. Один из них движется вглубь и соединяется с гипобластом вдоль его средней линии, оттесняя клетки гипобласта в стороны. Из этих движущихся в глубине клеток происходят все энтодермальные органы, а также большая часть внезародышевых оболочек (остальные формируются гипобластом). Второй мигрирующий поток распространяется по бластоцелю в виде рыхлого пласта примерно посередине между гипобластом и эпибластом. Этот пласт дает начало мезодермальным участкам зародыша и внезародышевых оболочек. К 22 ч инкубации основная часть презумптивных энтодермальных клеток оказывается внутри зародыша, тогда как презумптивные мезодермальные клетки еще продолжают мигрировать внутрь в течение длительного времени. Теперь начинается вторая фаза гаструляции. В то время как продолжается выселение клеток мезодермы, первичная полоска подвергается регрессии, в результате которой гензеновский узелок из своего положения вблизи центра area pellucida смещается к ее заднему краю (рис. 4.34). На этом пути он оставляет за собой дорсальную ось зародыша - головной отросток. По мере перемещения гензеновского узелка все далее кзади формируется остальной (задний)
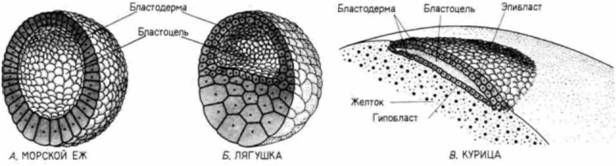
участок хорды. Наконец, узелок достигает своего окончательного положения на краю светлого поля, где и формирует анальную область зародыша на манер, свойственный истинно вторичноротым животным. К этому времени эпибласт состоит исключительно из презумптивных эктодермальных клеток В результате этого двухступенчатого процесса гаструляции зародыши птиц (и млекопитающих) обнаруживают отчетливый переднезадний градиент в зрелости их развития В то время как клетки задних участков зародыша еще претерпевают гаструляцию. клетки на его переднем конце уже начинают формировать органы. В течение нескольких следующих дней становится очевидным, что передний конец зародыша более продвинут в своем развитии, чем его задний коней. По мере того как презумптивные мезодермальные и энтодермальные клетки движутся внутрь, клетки-предшественники эктодермы окружают желток посредством эпиболии. Обрастание желтка эктодермой (снова напоминающее нам об эпиболии эктодермы у амфибий) задача для Геркулеса, осуществление которой занимает большую часть первых четырех дней инкубации и включает в себя непрерывное образование нового клеточного материала за счет запасов желтка, а также миграцию презумптивных эктодермальных клеток по нижней по- |