Гилберт С. Биология развития. Т.1.doc , БИР. Библиография Гилберт С. Биология развития в 3х т. Т. I Пер с англ. М. Мир, 1993. 228 с
 Скачать 10.9 Mb. Скачать 10.9 Mb.
|

Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с._______________ ГАСТРУЛЯЦИЯ: ПЕРЕРАСПРЕДЕЛЕНИЕ КЛЕТОК 125
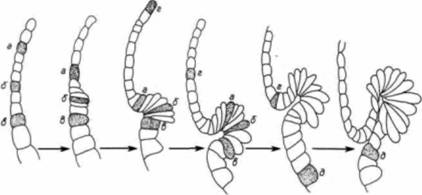
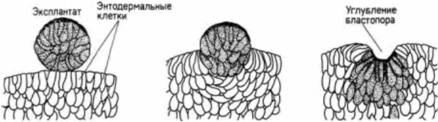
кладываться бластопор, лишь незадолго до начала гаструляции. Эти вегетативные клетки затем взаимодействуют с клетками, лежащими непосредственно над ними (в области серого серпа), побуждая их гаструлировать и формировать дорсальные осевые структуры. Клеточные движения и образование архентеронаГаструляция начинается с погружения группы краевых энтодермальных клеток, лежащих на спинной поверхности, внутрь зародыша. Это движение клеток внутрь осуществляется в результате изменения их адгезивных свойств, сходных с изменениями, которые были описаны ранее для клеток вегетативной пластинки у морских ежей. Рис. 4.21 иллюстрирует образование колбовидных клеток в ранней гаструле амфибий. Поскольку колбовидные клетки остаются в самой глубокой части бластопора и представляются тянущими за собой небольшой бластопор внутрь зародыша, долгое время считали, что именно они играют главную роль в формировании и увеличении размеров архентерона. Румблер (Rhumbler, 1902) предположил, что колбовидные клетки активно мигрируют в центρ гаструлы в результате низкого поверхностного натяжения. Они могут также втягивать, за собой новые соседние клетки, инициируя таким образом движение клеток внутрь зародыша. В 1943-1944 гг. выдающийся эмбриолог Иоганн Гольтфретер (Holtfreter, 1943, 1944) получил данные. свидетельствующие о том, что так и происходит на самом деле. Он изолировал клетки ранней губы бластопора и культивировал их на стекле. Округлившиеся концы клеток, прикрепленные к стеклу, вели за собой движение остальной части тела этих клеток. Еще более убедительными были рекомбинационные опыты Гольтфретера, в которых клетки краевой зоны (дающие начало спинной губе бластопора) комбинировали с клетками внутренней энтодермы. Когда клетки дорсальной краевой зоны изолировали и помещали на слой клеток внутренней эктодермы, из будущих клеток спинной губы бластопора формировались колбовидные клетки, которые погружались в эктодерму, опускаясь ниже ее поверхности (рис. 4.22). Кроме того, по мере погружения они создавали углубление, напоминающее бластопор в начале его образования. Таким образом, Гольтфретер доказал, что способность инвагинировать во внутреннюю энтодерму является прирожденным свойством клеток дорсальной краевой зоны. Сравнительно недавно Р. Келлер показал, что хотя клетки из области серого серпа действительно имеют внутренне присущую им способность инвагинировать в энтодерму, на самом деле колбовидные клетки играют менее активную роль, чем думали раньше. Келлер (Keller, I981 ) обнаружил, что после того, как сформировались колбовидные клетки, они не имеют особого отношения к увеличению размеров архентерона: частичное или полное удаление этих клеток не предотвращает инволюции прилежащих
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.126 ГЛАВА 4
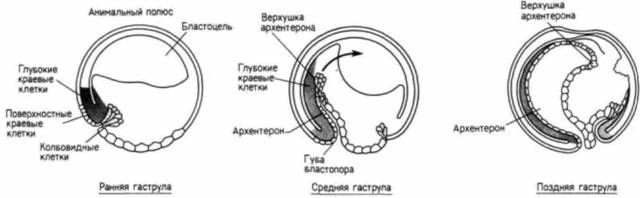
к ним клеток в бластопор. Гаструляция может происходить без колбовидных клеток. Главным фактором движения клеток внутрь зародыша является, вероятно, инволюция лежащих под его поверхностью краевых клеток, а не клеток, находящихся на поверхности. Эти лежащие под поверхностью глубокие инволюирующие краевые клетки вворачиваются внутрь и мигрируют к анимальному полюсу по внутренним поверхностям еще неинволюировавших глубоких клеток (рис. 4.23), а поверхностный слой формирует выстилку архентерона просто потому, что он прикреплен к активно мигрирующим глубоким клеткам. Даже если колбовидные клетки в какой-то степени участвуют в формировании первоначального углубления бластопора, движение этих клеток зависит от их прикрепления к лежащим под ними глубоким клеткам. В то время как удаление колбовидных клеток не влияет на инволюцию глубоких или поверхностных клеток краевой зоны внутрь зародыша, удаление глубоких клеток краевой зоны и их замещение клетками анимальной области (которые в норме не претерпевают инволюции) останавливает формирование архентерона. На рис. 4.24 делается попытка объединить результаты некоторых сравнительно недавних исследований (Keller, 1978, 1980, 1981; Keller, Schoenwolf, 1977) в виде модели, иллюстрирующей формирование архентерона. Согласно этой модели, во время начальных стадий гаструляции (рис. 4.24. А) краевая зона распространяется по направлению к вегетативному полюсу посредством интердигитации (вставления клеток из одного слоя в другой) нескольких слоев глубоких клеток, образующих в результате один тонкий слой. В то же самое время поверхностные клетки распространяются путем деления и уплощения. Морфология глубоких клеток, достигших бластопора, снова меняется, они формируют поток мезодермальных клеток, мигрирующих к анимальному полюсу по внутренней поверхности неинволюирующих глубоких клеток краевой зоны. В течение более поздних стадий гаструляции (рис. 4.24. Б) цилиндрические глубокие клетки уплощаются и растягиваются по мере того, как краевая зона продолжает смещаться в вентральном направлении (образуя широкий кольцевидный бластопор вокруг желточной пробки). Поток мезодермальных клеток продолжает мигрировать внутрь, и лежащий над ним слой поверхностных клеток (включая колбовидные) пассивно перемещается за ним в сторону анимального полюса, формируя энтодермальную крышу архентерона. Следовательно, хотя колбовидные клетки, возможно, и ответственны за образование начального углубления бластопора, однако движущая сила инволюции, по-видимому, заключена в глубоком слое краевых клеток. Кроме того, этот глубокий слой, вероятно, определяет продолжающуюся миграцию клеток внутрь зародыша. Миграция инволюирующей мезодермыМиграция клеток-предшественников мезодермы внутрь зародыша является частью удивительно хорошо координированной серии глобальных клеточных миграций. Подробная схема этих движений приведена на рис. 4.25. Один из наиболее важных аспектов гаструляции у амфибий это схождение к центру (конвергенция) и удлинение (растяжение) инволюирующей краевой зоны (ИКЗ), которая представляет собой область, расположенную непосредственно над губой бластопора. В поверхностном слое этой зоны (ИКЗП0В) находится материал энтодермальной крыши архентерона, а ее глубокий слой (ИКЗГЛ) содержит проспективные мезодермальные клетки (в том числе клетки хорды). |