Гилберт С. Биология развития. Т.1.doc , БИР. Библиография Гилберт С. Биология развития в 3х т. Т. I Пер с англ. М. Мир, 1993. 228 с
 Скачать 10.9 Mb. Скачать 10.9 Mb.
|

Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.ГАСТРУЛЯЦИЯ: ПЕРЕРАСПРЕДЕЛЕНИЕ КЛЕТОК 121
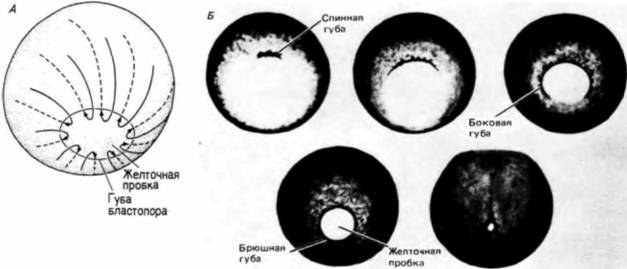
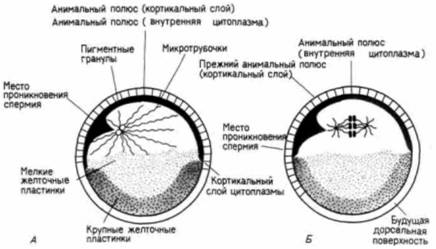
Судьба этих областей может быть предсказана еще до оплодотворения. Поверхность анимального полушария станет клетками эктодермы (эктодермальный эпителий и нервная система), поверхность вегетативного полушария даст клетки энтодермы (кишечный эпителий и связанные с ним органы), а мезодермальные клетки будут формироваться из внутренней цитоплазмы, лежащей вокруг экватора. Следовательно, зародышевые листки могут быть в общих чертах картированы в неоплодотворенном яйце: но это еще ничего не говорит нам о том, из какой части яйца образуется брюшная часть, а из какой - спинная. Переднезадняя (голова—хвост) и дорсовентральная (спина — живот) оси еще не детерминированы. Дорсовентральную и переднезаднюю оси детерминирует смещение цитоплазмы зиготы при оплодотворении. В гл. 2 мы обсуждали поворот кортикального слоя цитоплазмы относительно внутренней цитоплазмы яйца у лягушки. Внутренняя цитоплазма остается в прежнем положении из-за большого количества желтка в ней, тогда как кортикальный слой цитоплазмы активно смещается на 30о по отношению к своему первоначальному положению. Этот поворот приводит к тому, что анимально-вегетативная полярность наружного слоя яйца смещается на 30о по отношению к анимально-вегетативной полярности внутренней цитоплазмы. Таким образом возникает новое состояние симметрии. Вместо радиальной симметрии устанавливается двусторонняя, или билатеральная (рис. 4.17). Та область внутренней цитоплазмы вегетативного полушария, которая теперь лежит под кортикальным слоем цитоплазмы анимального полушария, по-видимому, активизируется и приобретает способность инициировать гаструляцию. Обычно это происходит на стороне, где образуется «серый серп», отстоящий на 180о от места проникновения спермия (рис. 4.17). Сторона, на которой спермий проникает в яйцо, станет брюшной поверхностью зародыша: противоположная сторона, где начинается гаструляция, будет спинной поверхностью зародыша (Gerhart et al.. 1981: Vincent et al., 1986; Gerhart et al., 1986). Для индукции этих движений цитоплазмы яйца спермий не является необходимым. Если искусственно стимулировать дробление энуклеированного яйца, то смещения кортикального слоя в положенное время все же происходит, однако направление его непредсказуемо. (В яйцах, оплодотворенных двумя спермиями, кортекс поворачивается только в одном направлении.) Спермий, по-видимому, дает сигнал для ориентации автономного смещения цитоплазмы в пространстве. Однако смещение цитоплазмы очень существенно: если оно блокировано, то дорсальные осевые структуры не формируются. Направление движения цитоплазмы определяет, какая сторона яйца должна стать спинной, а какая — брюшной (Gerhart et al., 1986; Vincent. Gcrhart, 1987). Направление смещения, определяемое местом проникновения спермия, может быть подавлено путем механического изменения пространственных со- Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.122 ГЛАВА 4
отношений между кортикальной и субкортикальной цитоплазмой. Это может быть достигнуто таким поворотом яйца, в результате которого внутренние компоненты попадают в разные области яйца (Pfluger. 1883; Penners. Schleip. 1928: Pasteels. 1948; Gerhart et al.. 1981). В норме оплодотворенное яйцо ориентировано таким образом, что тяжелые желточные пластинки в вегетативном полушарии в соответствии с действием силы тяжести располагаются в нижней части яйца. Если воспрепятствовать повороту яйца, погрузив его в раствор полисахарида (чтобы обезво-
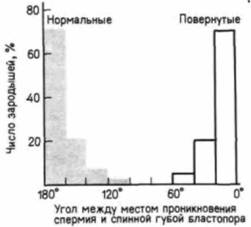
дить перивителлиновое пространство между яйцом и оболочкой оплодотворения), то затем яйцо можно повернуть набок на 90о, так чтобы анимально-вегетативная ось приняла вместо вертикального горизонтальное положение, а место проникновения спермия было обращено кверху (Gerhart et al.. 1981; Kirschncr, Gcrhart. 1981; Cooke. 1986). Если оплодотворенное яйцо остается в таком положении в течение 30 мин. начиная с середины первого цикла деления дробления, то почти у всех зародышей гаструляция начинается на той же стороне, на которой спермий проник в яйцо (рис. 4.18). Роль нормального смещения цитоплазмы можно продемонстрировать в опытах с подавлением этого смещения и последующим наблюдением за зародышами, развивавшимися из таких яиц. Воздействия, вызывающие деполимеризацию микротрубочек (холодовой шок, высокое гидростатическое давление или погружение яиц в нокодазол), блокируют поворот цитоплазмы (Mânes et al.. 1978). Обработанные таким образом яйца дробятся, но никогда не гаструлируют нормально и не приобретают дорсовентральной полярности. Гаструляция у них протекает симметрично, и они превращаются в трехслойный цилиндр, лишенный спинных компонентов, таких, как сомиты и центральная нервная система. Эти зародыши погибают после того, как будут исчерпаны их запасы желтка (Gerhart et al., 1986). Однако если эти яйца были повернуты на 90о за 40 мин до первого деления дробления (тем самым имитировался нормальный поворот цитоплазмы), то у зародышей формировались нормальные осевые структуры, а гаструляция начиналась на той стороне, которая должна была стать спинной (Sharf, Gerhart, 1983). Перераспределение цитоплазмы в течение первого цикла делений дробления является необходимым условием для нормальной гаструляции. Как и следовало ожидать из предыдущего |