Гилберт С. Биология развития. Т.1.doc , БИР. Библиография Гилберт С. Биология развития в 3х т. Т. I Пер с англ. М. Мир, 1993. 228 с
 Скачать 10.9 Mb. Скачать 10.9 Mb.
|

Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.112 ГЛАВА 4
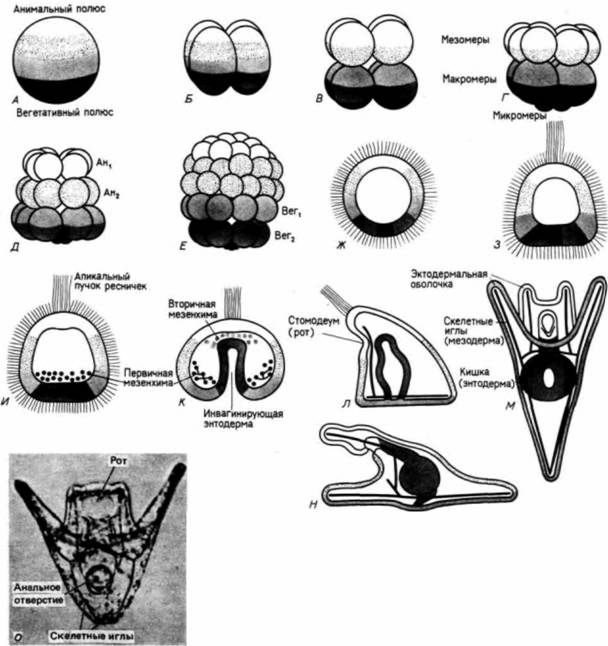
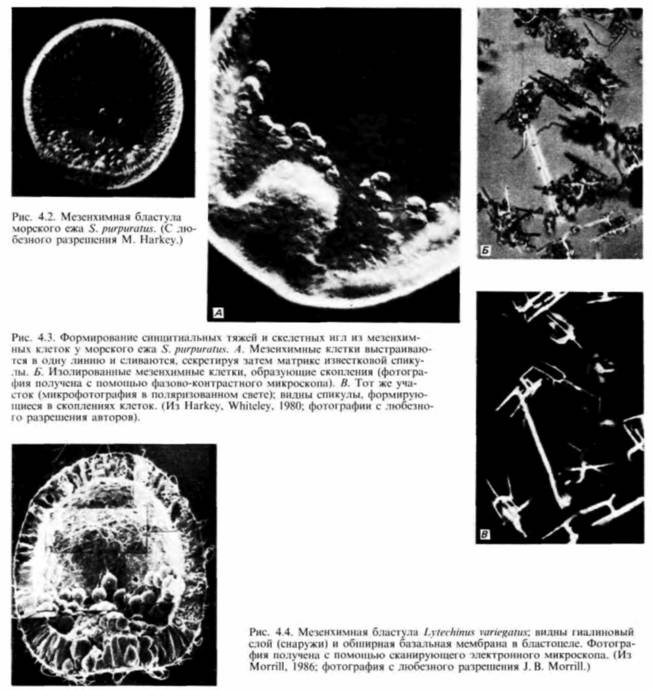
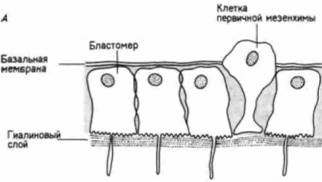
свойств по отношению к другим клеткам и окружающему их внеклеточному матриксу. Первоначально все клетки бластулы соединены по своей наружной поверхности с гиалиновым слоем (возникающим при оплодотворении из кортикальных гранул), а по внутренней — с базальной мембраной (внеклеточным матриксом, секретируемым клетками). Внеклеточный матрикс нельзя увидеть в световом микроскопе, но его можно обнаружить с помощью сканирующего электронного микроскопа (рис. 4.4). С боковых сторон каждая клетка имеет в качестве соседа другую клетку. Финк и Мак-Клей (Fink, McClay, 1985) подтвердили предположение Густафсона и Волперта. измерив силу адгезии бластомеров морского ежа с гиалиновым слоем, базальной мембраной и с другими клетками. Они обнаружили, что потомки макромеров и мезомеров очень прочно связаны друг с другом и с гиалиновым слоем, но Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.ГАСТРУЛЯЦИЯ ПЕРЕРАСПРЕДЕЛЕНИЕ КЛЕТОК 113
очень рыхло - с базальной мембраной (табл. 4.1). Характер связывания микромеров на стадии бластулы вначале был сходным. Однако в ходе гаструляции адгезивные свойства микромеров менялись. И если другие клетки сохраняли плотную связь с гиалиновым слоем и с соседними клетками, то микромеры гаструлы теряли свое сродство к этим структурам (оно уменьшалось до 2% первоначального значения), а сродство к базальной мембране увеличивалось в 100 раз. Эти изменения свойств микромеров приводят к тому, что они открепляются от наружного гиалинового слоя и от своих соседей и, притягиваемые базальной мембраной, мигрируют в бластоцель (рис. 4.5). Изменения в сродстве коррелировали Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.114 ГЛАВА 4
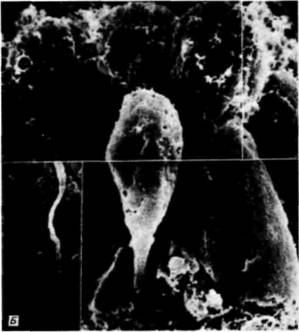
с происходившими в это время изменениями в составе молекул клеточной поверхности. Антиген клеточной поверхности, Мезо 1, обнаруживался точно тогда, когда наблюдалась миграция мезенхимных клеток из стенки бластулы (рисунок на внутренней стороне обложки). Как видно из рис. 4.4. вокруг вселяющихся в бластоцель клеток первичной мезенхимы концентрируется внеклеточный матрикс (Galileo. Morrill. 1985). Кроме того, попав внутрь бластоцеля, первичные мезенхимные клетки с помощью филоподий мигрируют по внеклеточному матриксу (рис. 4.6: Galileo. Morrill, 1985; Karp, Solursh, 1985). По-видимому, для их миграции важны два белка. Один из них — фибронектин, высокомолекулярный (400 000 дальтон) гликопротеин, который является обычным компонентом базальных мембран, в том числе и базальной мембраны у морского ежа (Wessel et al.. 1984). Финк и Мак-Клей (Kink. McCIay. 1985) показали, что во время гаструляции сродство микромеров к этому белку резко возрастает, а Като и Хайяши (Katoh. Hayashi. 1985) обнаружили, что миграция мезенхимных клеток зависит от концентрации фибронектина. Второй набор молекул — это сульфатированные гликопротеины, обнаруженные на клеточной поверхности выселяющихся в бластоцель мезенхимных клеток (Sugiyama. 1972; Heifctz, Lcnnarz, 1979). Если синтез (или сульфатирование) этих гликопротеинов подавлен, то мезенхимные клетки будут выселяться в бластоцель, но не станут двигаться в нем ( Karp. Solursh. 1974; Anstrom et al.. 1987; рис. 4.7). Гиббинс и др. (Gibbins et al.. 1969) показали, что для формирования и миграции мезенхимных клеток чрезвычайно важное значение имеет ориентация микротрубочек. В клетках ранней бластулы микротрубочки распределены асимметрично вдоль длинной оси клетки (рис. 4.8). По мере того как клетки первичной мезенхимы приобретают амебоидную форму и теряют контакт с бластодермой, микротрубочки |