Биохимия нервной ткани
 Скачать 91 Kb. Скачать 91 Kb.
|

|
Биохимия нервной ткани В.250599 БИОХИМИЯ НЕРВНОЙ ТКАНИ По своему соcтаву и процессам метаболизма нервная ткань значительно отличается от других тканей. Центральная функциональная клетка нервной ткани - нейрон - связана с помощью дендритов и аксонов с такими же клетками и клетками других типов, например, с секреторными и мышечными клетками. Клетки разделены синаптическими щелями. Связь между клетками осуществляется путем передачи сигнала. Сигнал проходит от тела нейрона по аксону до синапса. В синаптическую щель выделяется вещество-медиатор. Медиатор вступает в связь с рецепторами на другой стороне синаптической щели. Это обеспечивает восприятие сигнала и генерацию нового сигнала в клетке-акцепторе. ФУНКЦИИ НЕРВНОЙ ТКАНИ 1. Генерация электрического сигнала (нервного импульса) 2. Проведение нервного импульса 3. Запоминание и хранение информации. 4. Формирование эмоций и поведения. 5. Мышление. ОСОБЕННОСТИ ХИМИЧЕСКОГО СОСТАВА И МЕТАБОЛИЗМА НЕРВНОЙ ТКАНИ Специфику нервной ткани определяет гематоэнцефалический барьер (ГЭБ). ГЭБ имеет избирательную проницаемость для различных метаболитов, а также способствует накоплению некоторых веществ в нервной ткани. Например, в нервной ткани на долю глутамата и аспартата приходится примерно 70-75% от общего количества аминокислот. Таким образом, внутренняя среда нервной ткани намного отличается по химическому составу от других тканей. ЛИПИДЫ НЕРВНОЙ ТКАНИ. ФУНКЦИИ ЛИПИДОВ НЕРВНОЙ ТКАНИ. 1. Структурная: входят в состав клеточных мембран нейронов. 2. Функция диэлектриков (обеспечивают надежную электрическую изоляцию). 3. Защитная. Ганглиозиды являются очень активными антиоксидантами - ингибиторами перекисного окисления липидов (ПОЛ). При повреждении ткани мозга ганглиозиды способствуют ее заживлению. 4. Регуляторная. Фосфатидилинозиты являются предшественниками биологически активных веществ. Большая часть липидов нервной ткани находится в составе плазматических и субклеточных мембран нейронов и в миелиновых оболочках. В нервной ткани по сравнению с другими тканями организма содержание липидов очень высокое. Особенность липидного состава нервной ткани: есть фосфолипиды (ФЛ), гликолипиды (ГЛ) и холестерин (ХС), нет нейтральных жиров. Эфиры холестерина можно встретить только в участках активной миелинизации. Сам холестерин синтезируется интенсивно только в развивающемся мозге. В мозге взрослого человека низка активность ОМГ-КоА-редуктазы - ключевого фермента синтеза холестерина. Содержание свободных жирных кислот в мозге очень низкое. Некоторые нейромедиаторы после взаимодействия со специфическими рецепторами изменяют свою конформацию и изменяют конформацию фермента фосфолипазы С, которая катализирует расщепление связи в фосфатидилинозите между глицерином и остатком фосфата, в результате чего образуется фосфоинозитол и диацилглицерин. Эти вещества являются регуляторами внутриклеточного метаболизма. Диацилглицерин активирует протеинкиназу С, а фосфоинозитол вызывает повышение концентрации Са2+. Ионы кальция влияют на активность внутриклеточных ферментов и участвуют в работе сократительных элементов нервных клеток: микрофиламентов, что обеспечивает передвижение различных веществ в теле нервной клетки, аксоне и растущем кончике аксона. Протеинкиназа С участвует в реакциях фосфорилирования белков внутри нервных клеток. Если это белки-ферменты, то меняется их активность, если это рибосомальные или ядерные белки, то изменяется скорость биосинтеза белков. Липиды постоянно обновляются. Скорость их обновления различна, но в целом низка.. Некоторые липиды (например: холестерин, цереброзиды, фосфатидилэтаноламины, сфингомиелины) обмениваются медленно - в течение месяцев и даже лет. Исключение составляют фосфатидилхолин и, особенно, фосфатидилинозиты (содержат глицерин, фосфат, спирт (инозит), жирные кислоты) - они обмениваются очень быстро (сутки, недели). Синтез цереброзидов и ганглиозидов протекает с большой скоростью в развивающемся мозге в период миелинизации. У взрослых почти все цереброзиды (до 90%) находятся в миелиновых оболочках, а ганглиозиды - в нейронах. НУКЛЕИНОВЫЕ КИСЛОТЫ Нервные клетки не делятся, значит, не происходит синтез ДНК. Однако, содержание РНК в них самое высокое по сравнению с клетками остальных тканей организма. Скорость синтеза РНК тоже очень велика. В клетках нервной ткани не могут синтезироваться пиримидины (в нервной ткани отсутствует фермент карбамоилфосфатсинтаза). Пиримидины обязательно должны поступать из крови - ГЭБ для них проницаем. ГЭБ легко проницаем и для пуриновых мононуклеотидов, но, в отличие от пиримидиновых, они могут синтезироваться в нервной ткани. В нервной ткани, так же, как и в других, нуклеиновые кислоты обеспечивают хранение и передачу генетической информации и ее реализацию при синтезе клеточных белков. Например, сильные раздражители: громкие звуки, сильные зрительные стимулы и эмоции приводят к повышению скорости синтеза и РНК, и белка в определенных участках мозга. Это указывает на то, что изменения в нервной системе, отражающие индивидуальный опыт организма, кодируются в виде синтезированных макромолекул. Информация, благодаря которой нейроны устанавливают только определенные связи с определенными нейронами, кодируется в структуре полисахаридных веточек мембранных гликопротеинов. Образование таких связей, не заложенных в период эмбрионального развития, является результатом опыта индивидуального организма и составляет материальную основу для хранения информации, определяющей особенности поведения данного организма. МЕТАБОЛИЗМ УГЛЕВОДОВ И ОСОБЕННОСТИ ЭНЕРГЕТИЧЕСКОГО ОБЕСПЕЧЕНИЯ НЕРВНОЙ ТКАНИ. В нервной ткани, составляющей только 2% от массы тела человека, потребляется 20% кислорода, поступающего в организм. При этом энергетические возможности нервной ткани ограничены. 1. Основной путь получения энергии - только аэробный распад глюкозы по ГБФ-пути. Глюкоза является почти единственным энергетическим субстратом, поступающим в нервную ткань, который может быть использован ее клетками для образования АТФ. 2. Проникновение глюкозы в ткань мозга не зависит от действия инсулина, который не проникает через гематоэнцефалический барьер. Влияние инсулина проявляется лишь в периферических нервах. 3. Постоянный и непрерывный приток глюкозы и кислорода из кровеносного русла является необходимым условием энергетического обеспечения нервных клеток. Жесткая зависимость от поступления глюкозы обусловлена тем, что содержание гликогена в нервной ткани ничтожно (0.1% от массы мозга) и не может обеспечить мозг энергией даже на короткое время. С другой стороны, окисления неуглеводных субстратов с целью получения энергии не происходит. Поэтому при гипогликемии и/или даже кратковременной гипоксии в нервной ткани образуется мало АТФ. Следствием этого являются быстрое наступление коматозного состояния и необратимых изменений в ткани мозга. 4. Высокая скорость потребления глюкозы нервными клетками обеспечивается, в первую очередь, работой высокоактивной гексокиназы мозга. В отличие от других тканей, здесь гексокиназа не является ключевым ферментом всех путей метаболизма глюкозы. Гексокиназа мозга отличается низким значением Км и высокой Vmax, обладает в 20 раз большей активностью, чем соответствующий изофермент печени и мышц. Ключевыми ферментами ГБФ-пути в нервной ткани являются фосфофруктокиназа и изоцитратдегидрогеназа. Фосфофруктокиназу ингибируют фруктозо-1,6-бисфосфат, АТФ и цитрат, активируют фруктозо-6-фосфат, АДФ, АМФ и неорганический фосфат. Активность изоцитратДГ даже при нормальном уровне утилизации глюкозы в состоянии покоя максимальна. Поэтому при повышенном энергопотреблении нет возможностей ускорения реакций ЦТК. 5. Образование НАДФН2, который используется в нервной ткани в основном для синтеза жирных кислот и стероидов, обеспечивается сравнительно высокой скоростью протекания ГМФ-пути распада глюкозы. Энергия АТФ в нервной ткани используется неравномерно во времени. Так же, как и скелетные мышцы, функционирование нервной ткани сопровождается резкими перепадами в потреблении энергии. Резкое повышение энергозатрат происходит при очень быстром переходе от сна к бодрствованию. Поэтому существует еще одна особенность: 6. Образование креатинфосфата. Он обладает способностью удерживать макроэргические связи: 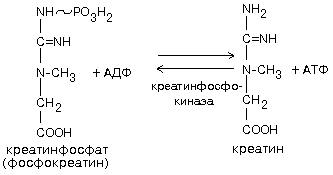 Эта реакция полностью обратима, ее направление зависит от соотношения АТФ/АДФ в клетках нервной ткани. Во время сна накапливается фосфокреатин. Переход к бодрствованию приводит к резкому уменьшению концентрации АТФ - равновесие реакции сдвигается влево, то есть образуется АТФ. МЕТАБОЛИЗМ АМИНОКИСЛОТ И БЕЛКОВ Ткань мозга интенсивно обменивается аминокислотами с кровью. Для этого существует специальные транспортные системы: две для незаряженных и еще несколько - для аминокислот, заряженных положительно и отрицательно. До 75% от общего количества аминокислот нервной ткани составляют аспартат, глутамат, а также продукты их превращений или вещества, синтезированные с их участием (глутамин, ацетильные производные, глутатион, ГАМК и другие). Их концентрации, и, в первую очередь, концентрация глутамата, в нервной ткани очень высоки. Например, концентрация глутаминовой кислоты может достигать 10ммоль/л. Функции глутамата в нервной ткани: 1. Энергетическая. Глутаминовая кислота связана большим числом реакций с промежуточными метаболитами ЦТК. 2. Глутамат (вместе с аспартатом) принимает участие в реакциях дезаминирования других аминокислот и временном обезвреживании аммиака. 3. Из глутамата образуется нейромедиатор ГАМК. 4. Глутамат принимает участие в синтезе глутатиона - одного из компонентов антиоксидантной системы организма. Глутаминовая кислота по праву занимает центральное место в обмене аминокислот мозга. Она используется для образования глутатиона, глутамина и гамма-аминомасляной кислоты. Образуется глутамат из своего кетоаналога - альфа-кетоглутаровой кислоты в ходе реакции трансаминирования. Реакция превращения альфа-КГ в глутамат протекает в ткани мозга с большой скоростью. Образующийся при этом глутамат являетя для ЦТК побочным продуктом. Большое расходование альфа-КГ восполняется за счет превращения аспарагиновой кислоты в метаболит ЦТК - ЩУК (см. рисунок). Образующаяся из глутамата ГАМК в результате нескольких реакций может быть превращена снова в ЩУК. Так образуется ГАМК-шунт, имеющийся в тканях головного и спинного мозга. Поэтому в этих тканях содержание ГАМК, как промежуточного метаболита циклического процесса, значительно выше, чем в остальных. На образование ГАМК здесь используется до 20% от общего количества глутамата. Остальные пути метаболизма аминокислот сходны с имеющимися в других тканях. До сих пор непонятным остается наличие в мозге почти полного набора ферментов орнитинового цикла, не содержащего карбамоилфосфатсинтазы, из-за чего мочевина здесь не образуется. 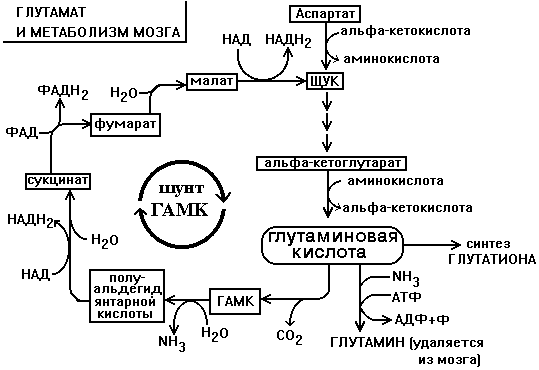 Ткань мозга способна синтезировать заменимые аминокислоты, как и другие ткани. НЕЙРОМЕДИАТОРЫ Нейромедиаторы - это вещества, которые характеризуются следующими признаками: 1. Накапливаются в пресинаптической структуре в достаточной концентрации; 2. Освобождаются при передаче импульса 3. Вызывают после связывания с постсинаптической мембраной изменение скорости метаболических процессов и возникновение электрического импульса. 4. Имеют систему для инактивации или транпортную систему для удаления из синапса, обладающие к ним высоким сродством. Таким образом, нейромедиаторы играют важную роль в функционировании нервной ткани, обеспечивая синаптическую передачу нервного импульса. Их синтез происходит в теле нейронов, а накопление - в особых везикулах, которые постепенно перемещаются с участием систем нейрофиламентов и нейротрубочек к кончикам аксонов. ХИМИЧЕСКАЯ КЛАССИФИКАЦИЯ НЕЙРОМЕДИАТОРОВ. 1) Аминокислоты (и их производные). К ним относят таурин, норадреналин, ДОФАминГАМК, глицин, ацетилхолин, гомоцистеин и некоторые другие (адреналин, серотонин, гистамин, серотонин). ТАУРИН. Таурин образуется из аминокислоты цистеина. Сначала происходит окисление серы в SH-группе до остатка серной кислоты (процесс идет в несколько стадий), а затем происходит декарбоксилирование. Таурин - это необычная кислота, в которой нет карбоксильной группы, а имеется остаток серной кислоты: 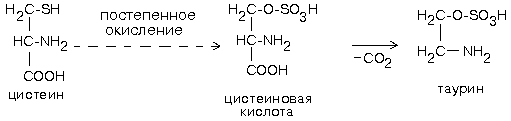 Таурин принимает участие в проведении нервного импульса в процессе зрительного восприятия. АЦЕТИЛХОЛИН Для синтеза холина требуются аминокислоты серин, метионин. Этаноламин может быть использован и в готовом виде. Но, как правило, из крови в нервную ткань пступает уже готовый холин. Второй же предшественник этого нейромедиатора - Ацетил-КоА, синтезируется в нервных окончаниях. 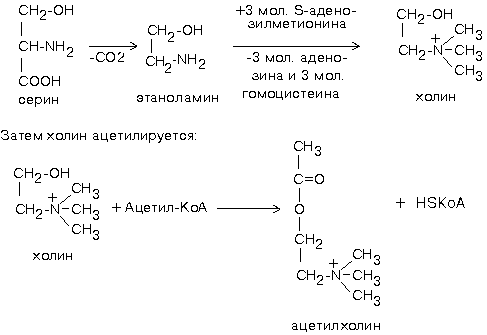 Продукт этой реакции ацетилхолин участвует в синаптической передаче нервного импульса. Он накапливается в синаптических пузырьках, образуя комплексы с отрицательно заряженным белком везикулином. Передача возбуждения с одной клетки на другую осуществляется с помощью специального синаптического механизма. Синапс - это функциональный контакт специализированных участков плазматических мембран двух возбудимых клеток. Синапс состоит из пресинаптической мембраны, синаптической щели и постинаптической мембраны. Мембраны клеток в месте контакта имеют утолщения в виде бляшек - нервных окончаний. Нервный импульс, достигший нервного окончания, не в состоянии преодолеть возникшее перед ним препятствие - синаптическую щель. После этого электрический сигнал преобразуется в химический. Пресинаптическая мембрана содержит специальные канальные белки, подобные белкам, формирующим натриевый канал в мембране аксона. Они тоже реагируют на мембранный потенциал, изменяя свою конформацию и формируют канал. В результате ионы Са2+ проходят через пресинаптическую мембрану по градиенту концентраций в нервное окончание. Градиент концентраций Са2+ создается работой Са2+-зависимой АТФазы - кальциевым насосом. Повышение концентрации Са2+ внутри нервного окончания вызывает слияние 200-300 имеющихся там везикул, заполненных ацетилхолином, с плазматической мембраной. Далее ацетилхолин секретируется в синаптическую щель путем экзоцитоза. и присоединяется к рецепторным белкам, расположенным на поверхности постсинаптической мембраны. Ацетилхолиновый рецептор представляет собой трансмембранный олигомерный гликопротеиновый комплекс, состоящий из 6 субъединиц:2-альфа, 2-бета, 1-гамма и 1-дельта. Плотность расположения белков-рецепторов в постсинаптической мембране очень велика - около 20000 молекул на 1 мкм2. Пространственная структура рецептора строго сооответствует конформации медиатора. При взаимодействии с ацетилхолином белок-рецептор так изменяет свою конформацию, что внутри него формируется натриевый канал. Катионная селективость канала обеспечивается тем, что ворота канала сформированы отрицательно заряженными аминокислотами. Таким образом повышается проницаемость постсинаптической мембраны для натрия и возникает новый импульс (или сокращение мышечного волокна). Деполяризация постсинаптической мембраны выеывает диссоциацию комплекса “ацетилхолин-белок-рецептор” и ацетилхолин освобождается в синаптическую щель. Как только ацетилхолин оказывается в синаптической щели, он за 40мкс подвергается быстрому гидролизу под действием фермента ацетилхолинэстеразы: 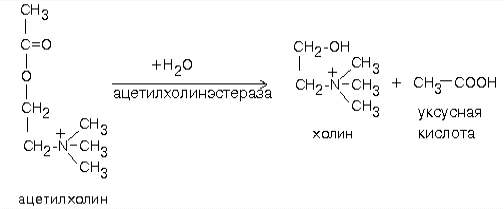 Во время гидролиза ацетилхолина образуется промежуточный фермент-субстратный комплекс, в котором ацетилхолин связан с активным центром фермента через серин. Необратимое ингибирование холинэстеразы вызывает смерть. Ингибиторами холинэстеразы являются фосфорорганические соединения (хлорофос, дихлофос, табун, зарин, зоман, бинарные яды). Эти вещества связываются ковалентно с серином в активном центре фермента. Некоторые из них синтезированы в качестве инсектицидов, а некоторые - в качестве БОВ (нервно-паралитические яды). Смерть наступает в результате остановки дыхания. Обратимые ингибиторы холинэстеразы используются как лечебные препараты. Например, при лечении глаукомы и атонии кишечника. КАТЕХОЛАМИНЫ: норадреналин и дофамин. Адренэргические синапсы встречаются в постганглионарных волокнах, в волокнах симпатической нервной системы, в различных отделах головного мозга. Катехоламины в нервной ткани синтезируются по общему механизму из тирозина. Ключевой фермент синтеза - тирозингидроксилаза, ингибируемая конечными продуктами. НОРАДРЕНАЛИН - медиатор в постганглионарных волокнах симпатической и в различных отделах ЦНС. ДОФАМИН - медиатор проводящих путей, тела нейронов которого расположены в отделе мозга, который отвечает за контроль произвольных движений. Поэтому при нарушении дофаминэргической передачи возникает заболевание паркинсонизм. Катехоламины, как и ацетилхолин, накапливаются в синаптических пузырьках и тоже выделяется в синаптическую щель при поступлении нервного импульса. Но регуляция в адренэргическом рецепторе происходит иначе. В пресинаптической мембране здесь имеется специальный регуляторный белок - альфа-ахромогранин (Мм=77 кДа), который в ответ на повышение концентрации медиатора в синаптической щели связывает уже выделившийся медиатор и прекращает его дальнейший экзоцитоз. Фермента, разрушающего медиатор, в адренэргических синапсах нет. После передачи импульса молекулы медиатора перекачивается специальной транспортной системой путем активного транспорта с участием АТФ обратно через пресинаптическую мембрану и включается вновь в везикулы. В пресинаптическом нервном окончании излишек медиатора может быть инактивирован МАО, а также катехоламин-О-метилтрансферазой путем метилирования по оксигруппе. Кокаин тормозит активный транспорт катехоламинов. Передача сигнала в адренэргических синапсах протекает по механизму, известному Вам из лекций по теме “Биохимия гормонов” с участием аденилатциклазной системы. Связывание медиатора с постсинаптическим рецептором почти мгновенно вызывает повышение концентрации ц-АМФ, что приводит к быстрому фосфорилированию белков постсинаптической мембраны. В результате изменяется генерация нервных импульсов постсинаптической мембраной (тормозится). В некторых случаях непосредственной причиной этого является повышение проницаемости постсинаптической мембраны для калия, либо снижением проводимости для натрия (эти события приводят к гиперполяризации). ГАМК ГАМК - тормозной медиатор. Повышает проницаемость постсинаптических мембран для ионов калия. Это ведет к изменению мембранного потенциала. ГЛИЦИН Тормозной медиатор, по вызываемым эффектам подобен ГАМК. 2) Пептиды Имеют в своем составе от трех до нескольких десятков аминокислотных остатков. Функционируют только в высших отделах нервной системы. Эти пептиды, как и катехоламины, выполняют функцию не только нейромедиаторов, но и гормонов. Передают информацию от клетки к клетке по системе циркуляции. Сюда относятся: а) Нейрогипофизарные гормоны (вазопрессин, либерины, статины). Эти вещества одновременно и гормоны и медиаторы. б) Гастроинтестинальные пептиды (гастрин, холецистокинин). Гастрин вызывает чувство голода, холецистокинин вызывает чувство насыщения, а также стимулирует сокращение желчного пузыря и функцию поджелудочной железы. в) Опиатоподобные пептиды (или пептиды обезболивания). Образуются путем реакций ограниченного протеолиза белка-предшественника проопиокортина. Взаимодействуют с теми же рецепторами, что и опиаты (например, морфин), тем самым имитируют их действие. Общее название - эндорфины - вызывают обезболивание. Они легко разрушаются протеиназами, поэтому их фармакологический эффект незначителен. г) Пептиды сна. Их молекулярная природа не установлена. Известно лишь, что их введение животным вызывает сон. д) Пептиды памяти (скотофобин). Накапливается в мозге крыс при тренировке на избегание темноты. е) Пептиды - компоненты РААС-системы. Показано, что введение ангиотензина-II в центр жажды головного мозга вызывает появление этого ощущения и стимулирует секрецию антидиуретического гормона. Образование пептидов происходит в результате реакций ограниченного протеолиза, разрушаются также под действием протеиназ. |