Через эндокринные железы и гормоны, которые секретируются в кровь транспортируются по кровеносному руслу и связываются с рецепторами клетокмишеней и влияют на метаболизм различных клетокмишеней
 Скачать 197.57 Kb. Скачать 197.57 Kb.
|

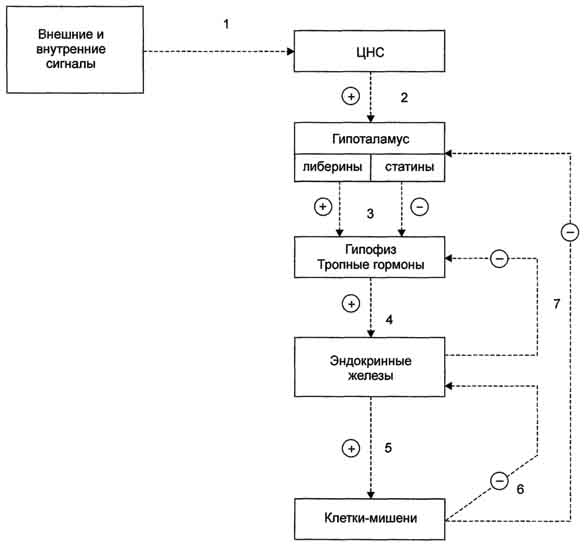
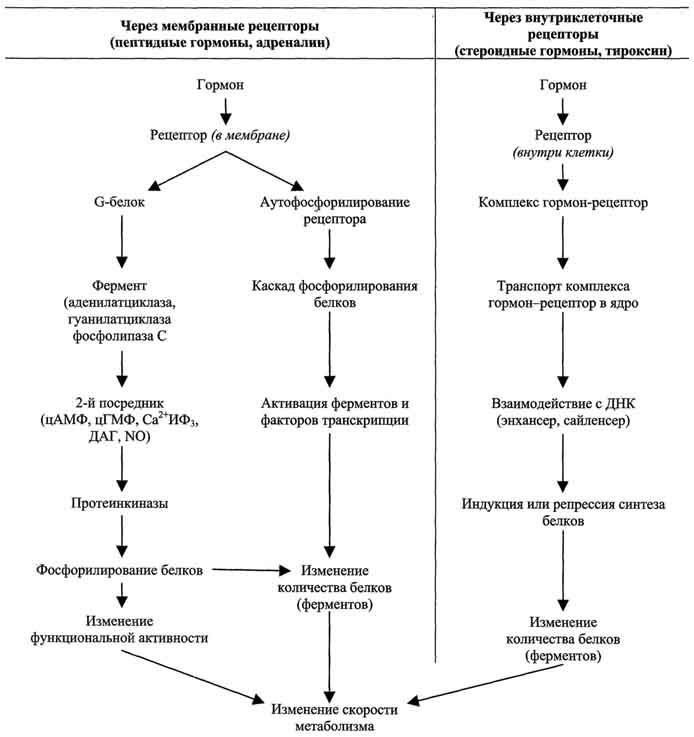
10-9 мол/л). Для образования комплекса центральные остатки соединительного района разматываются из альфа спирали чтобы образовать петлю (шарнир), который позволяет молекуле обернуться вокруг белка-мишени. N и C терминальные области сближаются друг с другом и их гидрофобные поверхности связываются с ним подобно двум рукам, удерживающим канат. Это способствует тому, что альфа спиральная последовательность мишени попадает в центр гидрофобного туннеля. Следствием этого является сильное изменение конформации белка-мишени. Как только концентрация Са2+ падает, комплекс диссоциирует, инактивируя белок. Однако есть и исключения. Так СаКМ-киназа II остается в активном состоянии после удаления Са2+. Тропонин С является изоформой КМ. Он присутствует в поперечно-полосатых мышцах, где регулирует взаимодействие между актином и миозином. 4 основные системы регуляции 4 основные системы регуляции Центральная и периферическая нервные системы через нервные импульсы и нейромедиаторы; Эндокринная система через эндокринные железы и гормоны, которые секретируются в кровь транспортируются по кровеносному руслу и связываются с рецепторами клеток-мишеней; и влияют на метаболизм различных клеток-мишеней; Паракринная и аутокринная системы посредством различных соединений, которые секретируются в межклеточное пространство и связываются с мембранными рецепторами соседних клеток. аутокринная - гормоны секретируются во внеклеточное пространство и связываются с мембранными рецепторами клетки, секретирующей гормон. Иммунная система через специфические белки (цитокины, антитела). изменения активности ферментов путём активации или ингибирования; изменения количества ферментов по механизму индукции или репрессии синтеза белков или изменения скорости их разрушения; изменения скорости транспорта веществ через мембраны клеток. Гуморальная регуляция — один из эволюционно ранних механизмов регуляции процессов жизнедеятельности в организме, осуществляемый через жидкие среды с помощью гормонов. гуморальная регуляция подчинена нервной регуляции и составляет совместно с ней единую систему нейрогуморальной регуляции. Продукты обмена веществ действуют не только непосредственно на эффекторные органы, но и на хеморецепторы и нервные центры, вызывая гуморальным или рефлекторным путём те или иные реакции. Поддержание уровня гормонов в организме обеспечивает механизм отрицательной обратной связи. Изменение концентрации метаболитов в клетках-мишенях по механизму отрицательной обратной связи подавляет синтез гормонов, действуя либо на эндокринные железы, либо на гипоталамус. Синтез и секреция тропных гормонов подавляется гормонами эндокринных периферических желёз. синтезируются в процессе иммунного ответа организма, служат медиаторами иммунной и воспалительной реакций и обладают в основном аутокринной, в некоторых случаях паракринной и эндокринной активностью; действуют как факторы роста и факторы дифференцировки клеток обладают плейотропной активностью. в результате изменения активности ферментов; в результате изменения количества ферментов связывание сигнальной молекулы, например гормона с рецептором (R), вызывает изменение конформации и увеличение сродства к Ор1с-белку. образование комплекса [Г] [К][Срlс-ГДФ] приводит к снижению сродства α-протомера Срlс-белка к ГДФ и увеличению сродства к ГТФ. ГДФ заменяется на ГТФ. это вызывает диссоциацию комплекса; отделившаяся α-субъединица, связанная с молекулой ГТФ, приобретает сродство к фос-фолипазе С. α-ГТФ взаимодействует с фосфолиггазой С и активирует её. Под действием фосфолипа-зы-С происходит гидролиз липида мембраны фосфатидилинозитол-4,5-бисфосфата (ФИФ2). в ходе гидролиза образуется и выходит в цитозоль гидрофильное вещество инозитол-1,4,5-трифосфат (ИФ3). Другой продукт реакции диацилглицерол (ДАГ) остаётся в мембране и участвует в активации фермента протеинкиназы С (ПКС). инозитол-1,4,5-трифосфат (ИФ3) связывается специфическими центрами Са2+-канала мембраны ЭР, это приводит к изменению конформации белка и открытию канала - Са2+ поступает в цитозоль. В отсутствие в цитозоле ИФ3 канал закрыт. Повышение концентрации Са2+ в цитозоле клетки увеличивает скорость взаимодействия Са2+ с неактивным цитозольным ферментом протеинкиназой С (ПКС) и белком кальмо-дулином, таким образом сигнал, принятый рецептором клетки, раздваивается. Связывание протеинкиназы С с ионами кальция позволяет ферменту вступать в кальций-опосредованное взаимодействие с молекулами "кислого" фосфолипида мембраны, фосфатидилсерина (ФС). Диацилглицерол, занимая специфические центры в протеинкиназе С, ещё более увеличивает её сродство к ионам кальция. На внутренней стороне мембраны образуется ферментативный комплекс - [ПКС][Са2+] [ДАГ][ФС] - активная протеинкиназа С, фосфорилирующая специфические ферменты по серину и треонину. 8 Естественными половыми стероидными гормонами являются эстрогены, андрогены и гестагены. По современным представлениям первой ступенью действия гормонов является их связывание со специфическими рецепторами в клетках органов-мишеней. Рецепторы гормонов стероидной структуры расположены в цитоплазме клеток, рецепторы гормонов пептидной и белковой структуры— на наружной мембране клетки-эффектора, гормоны щитовидной железы имеют свои специфические рецепторы в ядре клетки. Клетки-эффекторы имеют определенное число рецепторов. По химической структуре рецепторы относятся к белкам и выполняют две функции: распознают данный гормон среди других многочисленных молекул, соприкасающихся с клеткой; обеспечивают надлежащий клеточный ответ. Хотя для всех классов стероидных гормонов найдены специфические рецепторы, каждый стероидный гормон может реагировать с рецепторами других классов стероидов.. Механизм действия стероидных гормонов Проникновение стероида (С) в клетку Образование комплекса СР. Все Р стероидных гормонов представляют собой глобулярные белки примерно одинакового размера, с очень высоким сродством связывающие гормоны Трансформация СР в форму, способную связываться ядерными акцепторами [СР] Любая клетка содержит всю генетическую информацию. Однако при специализации клетки большая часть ДНК лишается возможности быть матрицей для синтеза иРНК. Это достигается путем сворачивания вокруг белков гистонов, что ведет к препятствию транскрипции. В связи с этим генетический материал клетки можно разделить на ДНК 3-х видов: 1.транскрипционно неактивная 2.постоянно экспрессируемая 3.индуцируемая гормонами или другими сигнальными молекулами. 9 Классификация гормонов и БАВ по химической структуре: Производные аминокислот: производные тирозина производные триптофана производные гистидина Белково-пептидные гормоны: полипептиды белки гликопротеиды Стероидные гормоны: кортикостероиды половые гормоны Производные жирных кислот: Функциональная классификация гормонов: Эффекторные гормоны — гормоны, которые оказывают влияние непосредственно на орган-мишень. Тропные гормоны — гормоны, основной функцией которых является регуляция синтеза и выделения эффекторных гормонов. Выделяются аденогипофизом. Рилизинг-гормоны — гормоны, регулирующие синтез и выделение гормонов аденогипофиза, преимущественно тропных. Выделяются нервными клетками гипоталамуса. Несмотря на то, что гормоны имеют разное химическое строение, для них характерны некоторые общие биологические свойства. 10 К настоящему времени в гипоталамусе открыто 7 стимуляторов (либерины) и 3 ингибитора (статины)секреции гормонов гипофиза: кортиколиберин, тиролиберин, люлиберин, фоллилиберин,соматолиберин, пролактолиберин, меланолиберин, соматостатин, пролактостатин и меланостатин. В чистом виде выделено 5 гормонов 1. Тиролиберин Тиролиберин представлен трипептидом. Нейромедиатор, повыш двиг активность, АД, стим секрецию тиреотроп гормонов 2. Гонадолиберин является декапептидом, состоящим из 10 аминокислот . Стимулирует секрецию ЛГ и ФСГ 3. Соматостатин является циклическим тетрадекапептидом. Вызывает торможение секреции тропных гормонов. 4. Соматолиберин Этот декапептид стимулирует синтез и секрецию гормона роста гипофиза соматотропина. 5. Кортиколиберин. Полипептид. Повыш синтез и секрецию адренокортикотропного гормона. 11 Гормоны передней доли гипофиза Гипофиз называют центральной железой, так как за счет его тропных гормонов регулируется деятельность других эндокринных желез. Гипофиз – состоит из аденогипофиза и нейрогипофиза К первой группе относят соматотропин и пролактин. соматотропин принимает участие в регуляции роста, усиливая образование белка. Наиболее выражено его влияние на рост эпифизарных хрящей конечностей. если имеется гиперфункция в детском возрасте, то развивается гигантизм; при гипофункции – карликовость. При гиперфункции у взрослого человека, увеличиваются те части тела, которые еще способны расти. Пролактин способствует образованию молока в альвеолах. После родов увеличивается синтез пролактина и наступает лактация. Ко второй группе гормонов относят: 1) тиреотропный гормон. Избирательно действует на щитовидную железу, повышает ее функцию. При сниженной выработке тиреотропина происходит атрофия щитовидной железы, при гиперпродукции – разрастание; 2) адренокортикотропный гормон. Стимулирует выработку глюкокортикоидов надпочечниками. Яв-ся антагонистом гормона роста. Он тормозит развитие основного вещества соединительной ткани, уменьшает количество тучных клеток, подавляет фермент гиалуронидазу, снижая проницаемость капилляров. Этим определяется его противовоспалительное действие. Секреция кортикотропина подвержена суточным колебаниям: в вечерние часы его содержание выше, чем утром; 3) гонадотропные гормоны а)фолликулостимулирующий гормон, стимулирующий рост и развитие фолликула в яичнике. Он незначительно влияет на выработку эстрагенов у женщин, у мужчин под его влиянием происходит образование сперматозоидов; б) лютеинизирующий гормон, стимулирующий рост и овуляцию фолликула с образованием желтого тела. Он стимулирует образование женских половых гормонов – эстрагенов, способствует выработке андрогенов у мужчин 12 Задняя доля гипофиза тесно связана с супраоптическим и паравентрикулярным ядром гипоталамуса. В нервных клетках паравентрикулярного ядра образуется окситоцин, в нейронах супраоптического ядра – вазопрессин. Вазопрессин: 1) усиливает сокращение гладких мышц сосудов; 2) угнетает образование мочи в почках. Антидиуретическое действие обеспечивается способностью вазопрессина усиливать обратное всасывание воды из канальцев почек в кровь. Окситоцин избирательно действует на гладкую мускулатуру матки, усиливает ее сокращение. Окситоцин стимулирует выделение молока. Акт сосания рефлекторно способствует выделению окситоцина из нейрогипофиза. Регуляция образования гормонов передней доли гипофиза осуществляется по принципу обратной связи. Между тропной функцией передней доли гипофиза и периферическими железами существуют двусторонние отношения: тропные гормоны активируют периферические эндокринные железы. Двусторонние взаимоотношения имеются между передней долей гипофиза и половыми железами, щитовидной железой и корой надпочечников. Тропные гормоны стимулируют функцию периферических желез, а гормоны периферических желез подавляют продукцию и выделение гормонов передней доли гипофиза. Повышение концентрации в крови гормона гипофиза приводит к торможению нейросекрета в гипоталамусе 13 В щитовидной железе синтезируются гормоны - йодтиронины. К ним относят 3,5,3'-трийодтиронин и 3,5,3',5'-тетрайодтиронин. Йодтиронины участвуют в регуляции многих процессов метаболизма, развития, клеточной дифференцировки, в регуляции экспрессии генов. Йодтиронины синтезируются в составе белка тиреоглобулина в фолликулах щитовидной железы. Тиреоглобулин синтезируется на рибосомах, далее поступает в аппарат Гольджи, а затем во внеклеточный коллоид, где он хранится и где происходит йодирование остатков тирозина. Образование йодтиронинов происходит в несколько этапов: транспорт йода в клетки щитовидной железы; окисление йода; йодирование остатков тирозина; образование йодтиронинов; транспорт йодтиронинов в кровь. Скорость синтеза и секреции йодтиронинов регулируются гипоталамо-гипофизарной системой по механизму обратной связи. Стимулом для повышения секреции тиреолиберина и тиреотропина служит снижение концентрации йодтиронинов в крови. Клетки-мишени йодтиронинов имеют 2 типа рецепторов к этим гормонам – 1. находятся в ядре участвуя в регуляции экспрессии генов. Другие рецепторы расположены в плазматической мембране клеток, обеспечивают связывание гормонов для удержания их у клетки. При физиологической концентрации йодтиронинов их действие проявляется в ускорении белкового синтеза, стимуляции процессов роста и клеточной дифференцировки. В печени йодтиронины ускоряют гликолиз, синтез холестерола и синтез жёлчных кислот. В печени и жировой ткани Т3 повышает чувствительность клеток к действию адреналина. В физиологических концентрациях Т3 увеличивает в мышцах потребление глюкозы, стимулирует синтез белков и увеличение мышечной массы. Гипотиреоз у новорождённых приводит к развитию кретинизма, задержкой умственного развития. Обычно гипотиреоз связан с недостаточностью функции щитовидной железы, но может возникать и при заболеваниях гипофиза и гипоталамуса. Наиболее тяжёлые формы гипотиреоза, сопровождающиеся слизистым отёком кожи и подкожной клетчатки- "микседема" . снижение частоты сердечных сокращений, вялость, сонливость, непереносимость холода, сухость кожи. Эти симптомы развиваются вследствие снижения основного обмена, скорости гликолиза, мобилизации гликогена и жиров, потребления глюкозы мышцами, уменьшения мышечной массы и снижения теплопродукции. Увзрослых людей частой причиной гипотиреоза является хронический аутоиммунный тиреоидит, приводящий к нарушению синтеза йодтиронинов (зоб Хашимото). Гипотиреоз может быть результатом недостаточного поступления йода в организм - эндемический зоб. Эндемический зоб часто встречается у людей, в районах, где содержание йода в воде и почве недостаточно. Если поступление йода в организм снижается (ниже 100 мкг/сут), то уменьшается продукция йодтиронинов, что приводит к усилению секреции ТТГ, под влиянием которого происходит компенсаторная гиперплазия щитовидной железы, но продукция йодтиронинов при этом не увеличивается. Гипертиреоз возникает вследствие повышенной продукции йодтиронинов. Диффузный токсический зоб (базедова болезнь) - наиболее распространённое заболевание щж. увеличение размеров щитовидной железы, повышение концентрации йодтиронинов в 2-5 раз и развитие тиреотоксикоза. увеличение основного обмена, учащение сердцебиений, мышечная слабость, снижение массы тела, потливость, повышение температуры тела, тремор и пучеглазие. 14. Гормоны паращитовидных желез. Химическая природа. Биологическая роль. Изменения метаболизма при гипо- и гиперпаратиреозе. Паратгормон синтезируется в паращитовидных железах и состоит из 84 аминокислотных остатков. Гормон хранится в секреторных гранулах. Секреция ПТГ регулируется уровнем кальция в крови: при снижении концентрации кальция происходит выделение гормона. Рецепторы для ПТГ расположены на поверхности клеток почек и костной ткани Эффекты гормона напралены на повышение концентрации ионов кальция и снижение концентрации фосфатов в крови. В почках ПТГ стимулирует реабсорбцию кальция в дистальных извитых канальцах, что ведет к уменьшению потерь кальция. Реабсорбция фосфатов наоборот уменьшается. ПТГ стимулирует образование кальцитриола из витамина Д3. Кальцитриол усиливает всасывание кальция в кишечнике. При связывании ПТГ с рецепторами клеток костной ткани их активность увеличивается: ускоряется образование щелочной фосфатазы, коллагеназы, которые вызывают распад костного матрикса. При этом происходит выход кальция и фосфатов из кости. Избыточная секреция ПТГ приводит к усилению реабсорбции кальция и выведения фосфатов, уровень кальция в крови увеличивается. Это проявляется мышечной слабостью, быстрой утомляемостью. Повышается риск возникновения переломов костей, образования камней в почках. Недостаточность паращитовидных желез проявляется гипокальциемией, повышением нервно-мышечной проводимости, судорожным синдромом. Кальцитонин – полипептид, состоящий из 32 аминокислот. Образуется в С-клетках паращитовидных желез и К-клетках щитовидной железы. По своему действию является антагонистом паратгормона. Клетками-мишенями для данного гормона являются клетки почек, костной ткани. Калицитонин снижает канальцевую реабсорцию кальция в почках. Активность остеокластов под действием кальцитонина снижается, что ведет к уменьшению распада костного матрикса. Скорость секреции кальцитонина зависит от уровня эстрогенов. При недостатке эстрогенов секреция кальцитонина снижается, что может привести к развитию остеопороза |