Онтогенез растительных клеток - StudentLib.com. Доклад На тему Онтогенез растительной клетки Игнатьев А. В. Проверил д б. н. Зайцева И. А
 Скачать 1.79 Mb. Скачать 1.79 Mb.
|

|
МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ,МОЛОДЕЖИ И СПОРТА УКРАИНЫ ДНЕПРОПЕТРОВСКИЙ НАЦИОНАЛЬНЫЙ УНИВЕРСИТЕТ им. ОЛЕСЯ ГОНЧАРА ФАКУЛЬТЕТ БИОЛОГИИ, ЭКОЛОГИИ И МЕДИЦИНЫ КАФЕДРА ФИЗИОЛОГИИ РАСТЕНИЙ Доклад На тему: Онтогенез растительной клетки Выполнил: Игнатьев А.В. Проверил: д.б.н. Зайцева И.А. Днепропетровск 2014 Содержание 1. Онтогенез растительной клетки . Эмбриональная фаза . Фаза растяжения . Фаза дифференцировки клетки . Фаза зрелости . Старение и смерть клетки Заключение 1. Онтогенез растительной клетки “Образовавшиеся в результате деления клетки проходят путь постепенного развития, который...” Онтогенез складывается из нескольких последовательных этапов - эмбриональной фазы, фазы роста, или растяжения, фазы дифференциации и, наконец, отмирания. Эмбриональные клетки отличаются небольшими размерами и содержат густую зернистую цитоплазму. Они располагаются плотно, без межклетников, имеют тонкую оболочку с преобладанием пектиновых веществ. 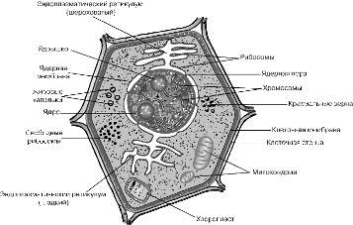 онтогенез растительная клетка В основе роста многоклеточных организмов лежит увеличение числа и размеров клеток, сопр6овождаемое их дифференциацией, т.е. возникновением и накоплением различий между клетками, образовавшимися в результате деления. Еще со времени Ю. Сакса рост клеток принято делить на три фазы: эмбриональную, растяжения, дифференцировки. Такое разделение носит условный характер. За последнее время внесены изменения в само понимание основных особенностей, характеризующих эти фазы роста. Если прежде считалось, что процесс деления клетки происходит лишь в эмбриональную фазу роста, то сейчас показано, что клетки могут иногда делиться и в фазу растяжения. Важно, что дифференцировка отнюдь не является особенностью только третьей, последней фазы роста. Дифференцировка клеток, в смысле появления и накопления внутренних физиологических различий между ними, проходит на протяжении всех трех фаз и является важной особенностью роста клеток. В третьей фазе эти внутренние физиологические различия лишь получают внешнее морфологическое выражение. Все же ряд существенных отличий между фазами роста имеется, и физиологи продолжают рассматривать их отдельно. Эмбриональная фаза. Клетка возникает в результате деления другой эмбриональной клетки. Затем она несколько увеличивается, главным образом за счет увеличения веществ цитоплазмы, достигает размеров материнской клетки и снова делится. Таким образом, эмбриональная фаза делится на два периода: период между делениями - интерфаза продолжительностью 15-20 ч и собственно деление клетки - 2-3 ч. Время это колеблется в зависимости от вида растений и условий (температуры). . Эмбриональная фаза Эмбриональная фаза или митотический цикл клетки делится на два периода: собственно деление клетки (2-3 ч) и период между делениями - интерфаза (15-20 ч). Митоз - это такой способ деления клеток, при котором число хромосом удваивается, так что каждая дочерняя клетка получает набор хромосом, равный набору хромосом материнской клетки. Структура клетки в интерфазе имеет ряд особенностей: густая цитоплазма с хорошо развитой эндоплазматической сетью, каналы которой узкие, с малым количеством расширений (цистерн); мелкие вакуоли; большое количество рибосом, многие из которых свободно располагаются в цитоплазме и не прикреплены к мембранам эндоплазматической сети; митохондрии много, но они еще не достигли окончательного размера, с малоразвитые кристами и густым матриксом. Имеются промитохондрии и пропластиды деление которых можно наблюдать. Ядро относительно небольшого размера, с крупным ядрышком. Нуклеоплазма - гомогенная, мелкозернистая. Хроматин в виде нитей и глыбок. Первичная клеточная оболочка тонкая, пронизана плазиодесмами. В период между делениями в клетке идут интенсивные процессы обмена веществ - активный синтез, высокая интенсивность дыхания, сопровождаемая образованием АТФ. Именно в этот период в ядре клетки происходит удвоение ДНК. Интерфаза, в свою очередь, делится на три триода: ) предсинтетический (g1), длительность 3-8 ч; ) синтетический (S) - 10-11 ч; ) постсинтетический (g2), - 4-5 ч. Переход из одного периода в другой регулируется ферментом циклинзависимой протеинкиназой. Большую роль в этом процессе играет белок циклин. При переходе от g1 -> S, g2 -> митоз происходит последовательное фосфорилирование и дефосфорилирование протеинкиназы, а также ее циклическое связывание с циклином и освобождение от него. В культуре тканей показано, что влияние фитогормонов на клеточный цикл осуществляется благодаря тому, что ауксин стимулирует синтез протеинкиназ, а цитокинин - циклина. В предсинтетический (g1) период ядро соматической клетки имеет определенное количество ДНК, характерное для данного вида организма. В этот период подготавливаются условия для репликации ДНК, происходит усиленный синтез РНК и соответствующих белков, в том числе белков-ферментов, участвующих в репликации. Сам процесс репликации начинается в синтетический период. Как известно, репликация ДНК - чрезвычайно сложный процесс, в котором участвует комплекс ферментов. Синтез РНК в период S, по-видимому, сокращается. Однако синтез белка продолжает идти, в частности образуются белки-гистоны. В постсинтетический период (перед переходом к митозу) в ядре уже находится четверное (соответствующее тетраплоидному) количество ДНК (материнские и дочерние молекулы), репликация ДНК прекращается, однако идет синтез РНК. Кроме того, происходит так называемый процессинг, при котором происходит превращение предшественника РНК (про-мРНК) в матричную РНК (мРНК). Репликация митохондриальной и пластидной ДНК происходит на протяжении всей интерфазы. Если процесс самовоспроизведения ДНК по какой-либо причине приостановлен, деление клетки не происходит. Таким образом, основные синтетические и энергетические процессы в клетке происходят именно в период между делениями. Существует несколько гипотез, объясняющих переход клетки к делению. Наиболее распространена гипотеза, согласно которой в меристематической клетке должно быть определенное соотношение между размерами ядра и цитоплазмы (ядерно-плазменное отношение). Когда это отношение ниже определенного уровня, ядро как бы уже не может управлять возросшей массой цитоплазмы и клетка переходит к делению. Перед делением происходят заметные изменения в энергетическом состоянии клетки. Во время интерфазы клетка характеризуется очень высоким энергетическим потенциалом - отношением АТФ к АМФ. При переходе к митозу благодаря глубокой структурной перестройке наступает как бы энергетическая разрядка и частично энергия выделяется в виде коротковолнового излучения (А.Г. Гурвич, В.Н. Жолкевич). В период деления интенсивность процессов обмена, в том числе и дыхания, падает. 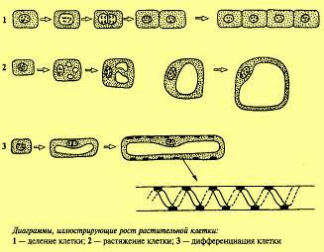 3. Фаза растяжения Делению клетки предшествует деление ядра. Первым признаком перехода ядра к делению является увеличение его объема. Это связано с возрастанием содержания воды и разжижением нуклеоплазмы. В ядре становятся видимыми извитые нити, сначала имеющие вид клубка. Постепенно нити утолщаются и укорачиваются. Выявляются хромосомы, каждая из которых состоит из двух того как клетка разделится 3-5 раз, она переходит во вторую фазу роста. Исключение составляют лишь инициальные клетки, которые продолжают делиться в течение всего периода роста растительного организма. Фаза растяжения. Переход к фазе растяжения сопровождается значительными структурными и физиологическими изменениями. Цитоплазма становится менее вязкой, более оводненной. Каналы эндоплазматической сети расширяются, в ряде мест они переходят в цистерны. Мембраны этой сети становятся шероховатыми, поскольку к ним прикрепляются рибосомы. Система внутренних мембран митохондрий (крист) получает полное развитие. Рост митохондриальных мембран происходит путем добавления новых компонентов. При этом белки и липиды синтезируются и включаются в мембраны координировано. Наблюдается увеличение контакта между митохондриями и эндоплазматической сетью, что облегчает снабжение энергией прикрепленных к ним рибосом. Ядро принимает неправильную форму, что увеличивает поверхность его соприкосновения с цитоплазмой. Размер ядрышка уменьшается. Мелкие вакуоли сливаются, и образуется одна центральная вакуоль. Увеличению объема вакуолей способствуют и присоединяющиеся к ним пузырьки, отделяющиеся от аппарата Гольджи. Доказано, что тонопласт образует инвагинации, благодаря чему в вакуоль попадают различные вещества цитоплазмы. Некоторые вещества и ферменты попадают в вакуоль с пузырьками, отделяющимися от аппарата Гольджи. В вакуолях накапливаются гидролитические ферменты, что способствует разложению различных органических веществ. В результате накопления ферментов, катализирующих распад крахмала, его содержание уменьшается и одновременно в вакуоли увеличивается количество Сахаров. Вместе с тем возрастает содержание аминокислот. Таким образом, в вакуоле заметно повышается содержание осмотически активных веществ. Относительное содержание цитоплазмы на единицу массы клетки падает, однако абсолютное ее содержание на клетку растет. Возрастает скорость синтеза белка, в клетке усиливаются все процессы метаболизма. Увеличение синтеза отдельных белков-ферментов происходит неравномерно. В результате меняется их соотношение, при этом в растягивающихся клетках разных органов это соотношение меняется по-разному, что и приводит к различным биохимическим и физиологическим особенностям (дифференциация). Высказывается предположение, что в фазу растяжения, по сравнению с фазой деления, не столько дерепрессируется работа новых генов, сколько в резкой степени активируется деятельность генов, уже разрешенных (Э.Е. Хавкин). При переходе к фазе растяжения заметно возрастает активность ферментов гликолиза и цикла Кребса. Увеличивается активность цитохромоксидазы. Интенсивность дыхания в расчете на клетку резко возрастает, меняется и качественная сторона дыхания. Повышается пропускная способность пентозофосфатного пути. В меристематических клетках скорость синтеза мРНК обгоняет синтез белка. В фазе растяжения, напротив, белок образуется в большем количестве по сравнению с РНК. Можно полагать, что в фазе растяжения синтез белка идет частично за счет пред образованных мРНК. Это подтверждается тем, что в меристематических клетках большая часть мРНК сосредоточена в ядре. В фазе растяжения количество мРНК в ядре уменьшается, а в цитоплазме растет. В результате изменения соотношения, а в определенной степени и набора ферментов в клетках в фазу растяжения меняются пути обмена веществ. В делящихся клетках мало протеолитических ферментов, в связи с этим распад белка заторможен. При переходе к растяжению активность протеолитических ферментов резко повышается, что способствует быстрому обновлению белков. Важно отметить также, что в меристематических клетках больше высокомолекулярных веществ по сравнению с низкомолекулярными (в расчете на клетку). В связи с этим у них не хватает субстратов и их метаболизм во многом зависит от соседних клеток. При переходе к растяжению возрастает количество низкомолекулярных веществ как в цитоплазме (активный фонд), так и в вакуоли, где они запасаются и при необходимости используются. Наиболее характерным процессом для фазы растяжения является значительное увеличение объема клетки. Скорость роста (в смысле увеличения объема) в эту фазу роста чрезвычайно велика. В течение всей фазы объем клетки возрастает в 20-50 и даже 100 раз. Это необратимое увеличение объема, идущее, главным образом, за счет усиленного поступления воды. Как известно, вода поступает в сторону меньшего (более отрицательного) водного потенциала. Водный потенциал - это мера активности воды, показывающая, во сколько раз активность (активность - это производное концентрации) воды в клетке меньше активности чистой воды. Водный потенциал Ψв клетки зависит от отрицательного осмотического потенциала и потенциала давления, который положителен: Ψв = Ψосм + Ψдавл. Вода поступает в сторону меньшего, более отрицательного водного потенциала. Частично уменьшение водного потенциала и, как следствие, возрастание поступления воды может идти за счет возрастания концентрации осмотически действующих веществ. Как уже упоминалось, этому способствует усиление активности гидролитических ферментов в вакуолярном соке. Однако поступление воды идет значительно более интенсивно, чем это может быть объяснено исходя из изменения осмотической концентрации. Основной причиной является, по-видимому, уменьшение противодавления клеточной оболочки (потенциала давления) или ее рост. Как уже рассматривалось, первичная клеточная оболочка построена таким образом, что, с одной стороны, она может противостоять гидростатическому давлению, развивающемуся внутри клетки, а с другой - способна к растяжению эластическому (обратимому) и пластическому (необратимому). Эти свойства обеспечиваются тем, что состав клеточной оболочки составляют волокна - микрофибриллы, которые погружены в пластический матрикс. Ориентация микрофибрилл целлюлозы не безразлична. В первичной клеточной оболочке в период растяжения микрофибриллы располагаются либо беспорядочно, либо в основном перпендикулярно продольной оси клетки. Последнее способствует удлинению клетки под влиянием гидростатического (тургорного) давления. Важно отметить, что рост растяжением начинается тогда, когда тургор достигает пороговой величины и дальнейшее поступление воды возможно лишь за счет пластического (необратимого) растяжения клеточной оболочки. Большую роль в ориентации микрофибрилл играют микротрубочки. В растущих клетках микротрубочки, расположенные около клеточной стенки, также ориентированы перпендикулярно оси растяжения. Поскольку микрофибриллы целлюлозы не растягиваются в длину, то при растяжении они скользят друг около друга. Как видно из приведенной выше схемы, микрофибриллы и вещества матрикса оболочки связаны между собой различными связями. В период роста растяжением эти связи (водородные и ковалентные) разрыхляются. Но после того как растяжение произошло, связи вновь возникают. Разрыв водородных связей может регулироваться различными внешними условиями. Одним из таких условий может быть концентрация ионов водорода (рН). Известно, что водородные связи ослабевают при низких значениях рН. В клеточных оболочках локализован ряд ферментов, по преимуществу гидролаз. Эти ферменты, расщепляя соответствующие связи, могут участвовать в растяжении клеточной оболочки. Особенное значение имеет, по-видимому, 1,3-глюконазы-ферменты, расщепляющие связи между остатками молекулы глюкозы. Интересно отметить, что активность гидролаз, локализованных в клеточных оболочках, повышается при подкислении. Таким образом, снижение значения рН способствует разрыву связей как водородных, так и ковалентных между компонентами клеточной оболочки, ее разрыхлению. Как следствие клеточная оболочка становится более рыхлой, ее сопротивление (противодавление) падает и становится возможным дополнительное поступление воды. Большую роль в этом процессе играет фитогормон ауксин, который регулирует рост растяжением. Рост клеточной оболочки связан с новообразованием ее составляющих. Подобно тому, как указывалось при рассмотрении новообразования клеточной оболочки при делении клеток, пектиновые вещества образуются из галактуроновой кислоты в везикулах аппарата Гольджи. Везикулы подходят к плазмалемме, и их мембраны сливаются с ней, а нецеллюлозные компоненты включаются в клеточную оболочку. Микрофибриллы целлюлозы синтезируются на внутренней поверхности клеточной оболочки, прилегающей к плазмалемме, из веществ цитоплазмы с помощью ферментов, поставляемых из аппарата Гольджи. В образовании клеточной стенки принимает участие и ЭПР В шероховатом ЭПР синтезируется белок экстенсии. Существует несколько типов роста клеточной оболочки: ) вновь образовавшиеся микрофибриллы целлюлозы внедряются в промежутки между сетью старых микрофибрилл (интусессцепция); ) сетка вновь образовавшихся микрофибрилл целлюлозы, между которыми образуются новые связи, накладывается на старую. При этом происходит и переориентировка старых молекул: они становятся в более вертикальное положение. Общая толщина стенки при этом не изменяется, оставаясь около 0,3-0,5 мкм. Этот особенный тип аппозиционного роста получил название многосетчатого роста. Таким образом, рост растяжением включает следующие этапы: ) разрыхление связей между компонентами клеточной оболочки и увеличение ее пластичности; ) поступление воды, которая давит на стенки, вызывает растяжение и увеличивает объем клетки; ) закрепление увеличения объема путем многосетчатого роста оболочки. В конце фазы растяжения усиливается лигнификация клеточных стенок, что снижает ее упругость и проницаемость, накапливаются ингибиторы роста, повышается активность оксидазы ИУК, снижающей содержание ауксина в клетке. . Фаза дифференцировки клетки Фаза дифференцировки клетки. Каждая клетка растения содержит в своем геноме полную информацию о развитии всего организма и может дать начало формированию целого растения (свойство тотипотентности). Однако, находясь в составе организма, эта клетка будет реализовать только часть своей генетической информации. Сигналами для экспрессии только определенных генов служат сочетания фитогормонов, метаболитов и физико-химических факторов (например, давление соседних клеток). На этой фазе процесс дифференцировки уже проявляется в определенных структурных признаках, т. е. меняется форма, внутренняя и внешняя структура клетки. Процесс функциональной дифференциации клеток, или накопление физиологических различий между ними, происходит на всех фазах роста. Определенные различия имеются уже между появившимися в период деления дочерними клетками, из которых в дальнейшем будут образовываться различные ткани. Это проявляется в их химическом составе, морфологических особенностях. Значительно варьируют число и структура митохондрий, и особенно пластид, обилие и локализация эндоплазматической сети. Очень видоизменяются клетки проводящей системы. При дифференциации члеников ситовидных трубок большинство органелл разрушается. В сосудах ксилемы почти полностью исчезает цитоплазма. Происходит образование вторичной клеточной оболочки. Этот процесс сопровождается наложением новых слоев микрофибрилл целлюлозы на старые. При этом ориентация фибрилл целлюлозы в каждом новом слое другая. Клеточная оболочка утолщается и теряет способность к росту. . Фаза зрелости  Фаза зрелости. Клетка выполняет те функции, которые заложены в ходе ее дифференцировки. . Старение и смерть клетки Старение и смерть клетки. При старении клеток происходит ослабление синте-тических и усиление гидролитических процессов. В органеллах и цитоплазме образуются автофагические вакуоли, разрушаются хлорофилл и хлоропласты, эндоплазматический ретикулум, аппарат Гольджи, ядрышко, набухают митохондрии, в них снижается число крист, вакуолизируется ядро. Гибель клетки становится необратимой после разрушения клеточных мембран, в том числе и тонопласта, выхода содержимого вакуоли и лизосом в цитоплазму. Старение и смерть клетки происходит в результате накопления повреждений в генетическом аппарате, клеточных мембранах и включения генетической програмированной клеточной смерти - PCD (programmed cell death), аналогичной апоптозу у клеток животных. Заключение Под термином Онтогенез (греч. On - род. падеж, Ontos - Существо; лат. GeneSis - происхождение, формирование) понимают индивидуальное развитие организма, органа, клетки от момента образования до естественной гибели. Онтогенез многих растительных клеток проходит по первому варианту - от митоза до естественной смерти. В этом случае он также слагается из нескольких этапов: деление клетки, рост растяжением, дифференцировка, активное функционирование, старение и смерть. В процессе дифференцировки формируются различия между клетками, т. е. происходит их специализация. Деление на этапы достаточно условно: дифференцировка клетки, в частности, начинается уже на стадии роста. Особенностью растительных клеток является возможность ее дедифференцирования, т. е. перехода специализированных клеток обратно к пролиферации. Это происходит при образовании каллуса (травматической ткани двудольных), а также In Vitro При получении культур клеток. Культивируемые клетки можно получить из любой ткани любого органа растения, т. е. к дедифференциации способна практически любая живая клетка растения, независимо от типа ее дифференцировки. |