Экология учебник для вузов - А.С. Степановских. Экология
 Скачать 21.33 Mb. Скачать 21.33 Mb.
|

|
Физиологические адаптации растений к световым условиям наземно-воздушной среды охватывают различные жизненные функции. Установлено, что у светолюбивых растений ростовые процессы более чутко реагируют на недостаток света по сравнению с теневыми. В результате наблюдается усиленное вытягивание стеблей, которое помогает растениям пробиться к свету, в верхние ярусы растительных сообществ. Основные физиологические адаптации к свету лежат в сфере фотосинтеза. В общей форме изменение фотосинтеза в зависимости от интенсивности света выражается «световой кривой фотосинтеза». Экологическое значение имеют следующие ее параметры (рис. 5.23). 1. Точке пересечения кривой с осью ординат (рис. 5.23, а) соответствует величина и направление газообмена растений в полной темноте: фотосинтез отсутствует, имеет место дыхание (не поглощение, а выделение СО2), поэтому точка а лежит ниже оси абсцисс. 2. Точка пересечения световой кривой с осью абсцисс (рис. 5.23, б) характеризует «компенсационный пункт», т. е. интенсивность света, при которой фотосинтез (поглощение СО2) уравновешивает дыхание (выделение СО2). 3. Интенсивность фотосинтеза с увеличением света возрастает только до определенного предела, в дальнейшем остается постоянной — световая кривая фотосинтеза выходит на «плато насыщения». 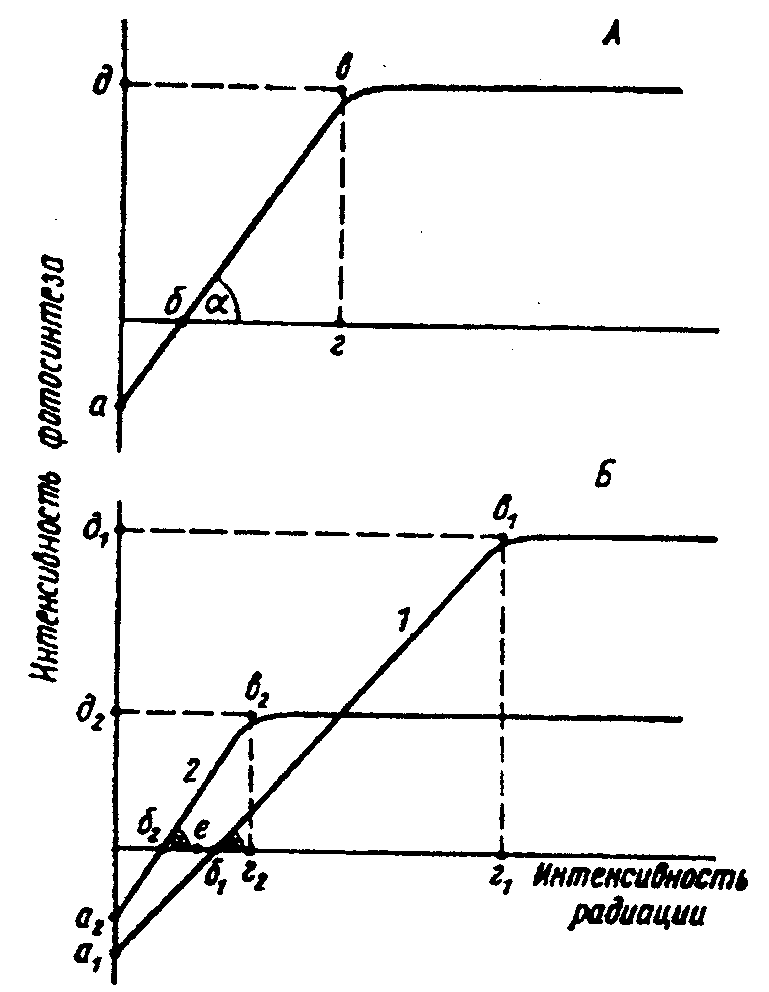 Рис. 5.23. Световые кривые фотосинтеза: А — общая схема; Б — кривые для светолюбивых (1) и теневыносливых (2) растений На рис. 5.23 область перегиба условно обозначена плавной кривой, перелому которой соответствует точка в. Проекция точки в на ось абсцисс (точка г) характеризует «насыщенную» интенсивность света, т. е. такую величину, выше которой свет уже не повышает интенсивность фотосинтеза. Проекция на ось ординат (точка д) соответствует наибольшей интенсивности фотосинтеза для данного вида в данной наземно-воздушной среде. 4. Важная характеристика световой кривой — угол наклона (а) к абсциссе, которая отражает степень увеличения фотосинтеза при возрастании радиации (в области сравнительно низкой интенсивности света). У растений отмечается сезонная динамика реакции на свет. Так, у осоки волосистой (Carex pilosa) ранней весной в лесу только что появившиеся листья имеют плато светового насыщения фотосинтеза за 20 — 25 тыс. лк, при летнем затенении у этих же видов кривые зависимости фотосинтеза от света становятся соответственными параметрам «теневым», т. е. листья приобретают способность более эффективно использовать слабый свет, эти же листья после перезимовки под пологом безлистного весеннего леса снова обнаруживают «световые» черты фотосинтеза. Своеобразной формой физиологической адаптации при резком недостатке света служит потеря растением способности к фотосинтезу, переход к гетеротрофному питанию готовыми органическими веществами. Иногда такой переход становился безвозвратным из-за потери растениями хлорофилла, например, орхидеи тенистых еловых лесов (Goodyera repens, Weottia nidus avis), вертляница (Monotropa hypopitys). Они живут за счет мертвых органических остатков, получаемых от древесных пород и других растений. Данный способ питания получил название сапрофитного, а растения называют сапрофитами. Для подавляющего большинства наземных животных с дневной и ночной активностью зрение представляет один из способов ориентации, имеет важное значение для поисков добычи. Многие виды животных обладают и цветным видением. В связи с этим у животных, особенно жертв, возникли приспособительные особенности. К ним относятся защитная, маскирующая и предупреждающая окраска, покровительственное сходство, мимикрия и т. п. Возникновение ярко окрашенных цветков высших растений также связано с особенностями зрительного аппарата опылителей и в конечном счете со световым режимом среды. Водный режим. Дефицит влаги — одна из наиболее существенных особенностей наземно-воздушной среды жизни. Эволюция наземных организмов проходила путем приспособления к добыванию и сохранению влаги. Режимы влажности среды на суше разнообразны — от полного и постоянного насыщения воздуха водяными парами, где в год выпадает несколько тысяч миллиметров осадков (области экваториального и муссонно-тропического климата) до практически полного их отсутствия в сухом воздухе пустынь. Так, в тропических пустынях среднегодовое количество осадков меньше 100 мм в год, и при этом дожди выпадают не каждый год. Годовое количество осадков не всегда дает возможность оценить водообеспеченность организмов, так как одно и то же их количество может характеризовать пустынный климат (в субтропиках) и очень влажный (в Арктике). Большую роль играет соотношение осадков и испаряемости (суммарного годового испарения со свободной водной поверхности), также неодинаковый в разных районах земного шара. Области, где эта величина превышает годовую сумму осадков, называют аридными (сухими, засушливыми). Здесь, например, растения испытывают недостаток влаги в течение большей части вегетационного периода. Области, в которых растения обеспечены влагой, называют гумидными, или влажными. Нередко выделяют и переходные зоны — полуаридные (семиаридные). Зависимость растительности от среднегодового количества осадков и температуры показана на рис. 5.24. 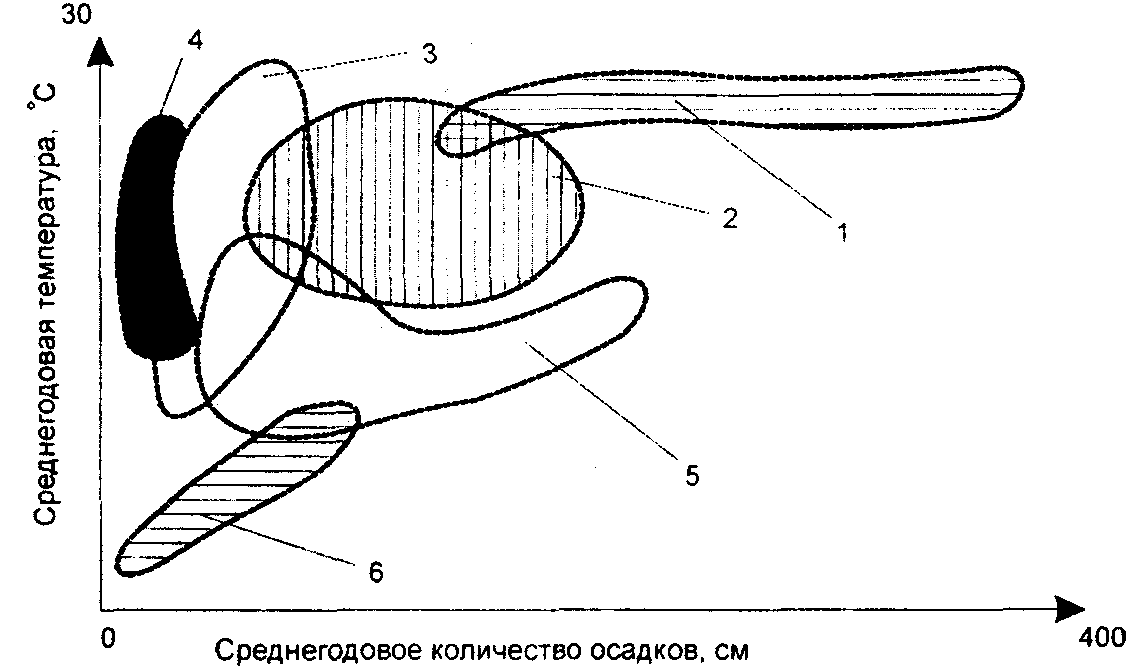 Рис. 5.24. Зависимость растительности от среднегодового количества осадков и температуры: 1 — тропический лес; 2 — листопадный лес; 3 — степь; 4 — пустыня; 5 — хвойный лес; 6 — арктическая и горная тундра Водообеспечение наземных организмов зависит от режима выпадения осадков, наличия водоемов, запасов почвенной влаги, близости грунтовых вод и т. д. Это способствовало развитию у наземных организмов множества адаптации к различным режимам водообеспечения. На рис. 5.25 слева направо показаны переход от обитающих в воде низших водорослей с клетками без вакуолей к первичным пойкилогидрическим наземным водорослям, образование вакуолей у водных зеленых и харовых водорослей, переход от имеющих вакуоли таллофитов к гомойогидрическим кормофитам (распространение мхов — гидрофитов еще ограничено местообита-ниями с высокой влажностью воздуха, в сухих местообитаниях мхи становятся вторично пойкилогидрическими); среди папоротников и покрытосеменных (но не среди голосеменных) также имеются вторично пойкилогидрические формы. Большинство ли-стостебельных растений гомойогидричны благодаря наличию у них кутикулярной защиты от транспирации и сильной вакуолизации их клеток. Следует отметить, что ксерофильность животных и растений свойственна только наземно-воздушной среде. 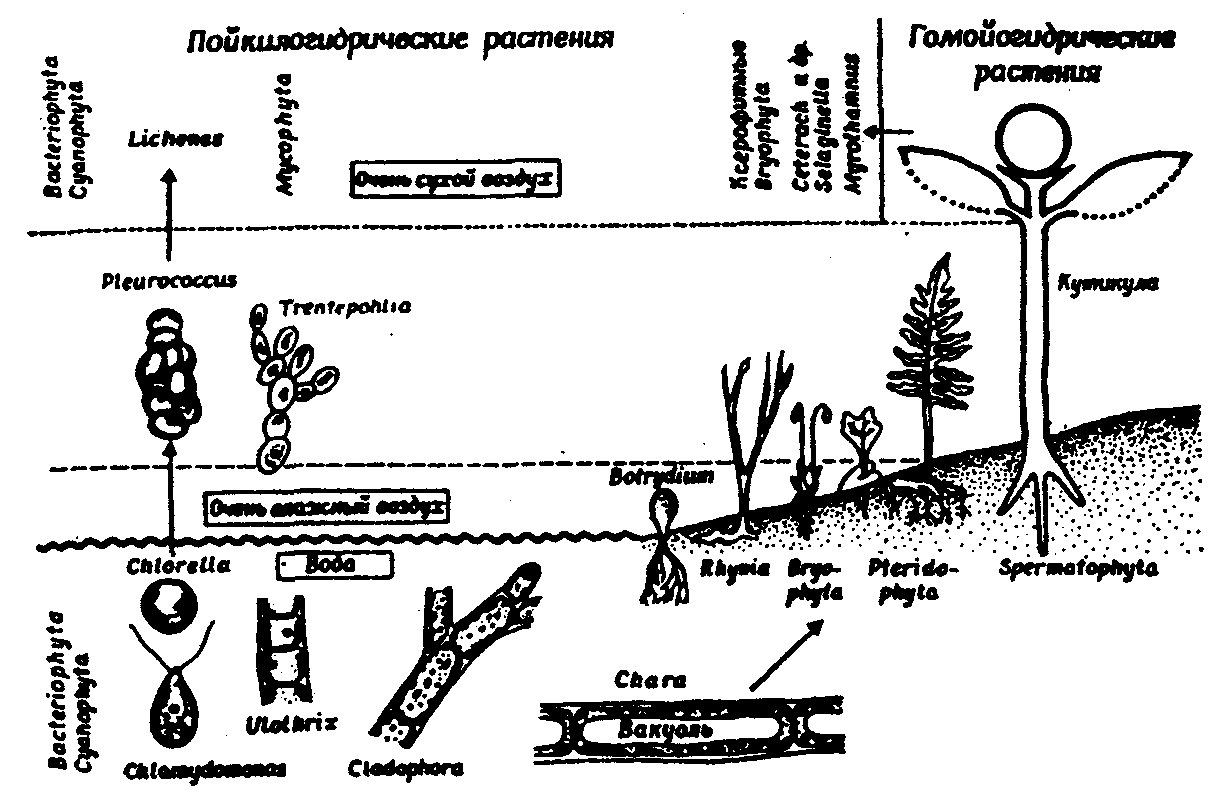 Рис. 5.25. Приспособление водного обмена растений к наземному образу жизни (из В. Лархера, 1978) Осадки (дождь, град, снег), кроме водообеспечения и создания запасов влаги, часто играют и другую экологическую роль. Например, при ливневых дождях почва не успевает впитывать влагу, вода сильными потоками быстро стекает и зачастую сносит в озера и реки слабо укоренившиеся растения, мелких животных и плодородный слой почвы. В поймах рек дожди могут вызывать паводки и оказывать таким образом неблагоприятное воздействие на обитающих здесь растения и животных. В затопляемых периодически местах образуются своеобразные пойменные фауна и флора. Отрицательное действие на растения и животных оказывает и град. Посевы сельскохозяйственных культур на отдельных полях иногда бывают полностью уничтожены этим стихийным бедствием. Многообразна экологическая роль снежного покрова. Для растений, почки возобновления которых находятся в почве или у ее поверхности, многих мелких животных снег играет роль теплоизо-лирующего покрова, защищая от низких зимних температур. При морозах выше -14°С под слоем снега 20 см температура почвы не опускается ниже 0,2°С. Глубокий снежный покров предохраняет от вымерзания зеленые части растений, такие, как вероника лекарственная, копытень и др., которые уходят под снег, не сбрасывая листвы. Мелкие наземные животные ведут зимой активный образ жизни, прокладывая под снегом и в его толще многочисленные галереи ходов. При наличии витаминизированного корма в снежные зимы там могут размножаться грызуны (лесная и желтогорлая мыши, ряд полевок, водяная крыса и др.). Под снегом в сильные морозы прячутся рябчики, куропатки, тетерева. Крупным животным зимний снежный покров нередко мешает добывать корм, передвигаться, особенно при образовании на поверхности ледяной корки. Так, лоси (Alces alces) свободно преодолевают слой снега глубиной до 50 см, но более мелким животным это недоступно. Часто при многоснежных зимах наблюдается гибель косуль, диких кабанов. Выпадение большого количества снега оказывает отрицательное влияние и на растения. Помимо механических повреждений в виде снеголомов или снеговалов мощный слой снега может приводить к выпреванию растений, а во время таяния снега, особенно в затяжную весну, к вымоканию растений. Рис. 5.26. Опорная поверхность конечностей белой куропатки зимой (А) и летом (Б) О  т низких температур при сильных ветрах в малоснежные зимы страдают растения и животные. Так, в годы, когда снега выпадает мало, гибнут мышевидные грызуны, кроты и другие мелкие животные. Вместе с тем в широтах, где зимой выпадают осадки в виде снега, растения и животные исторически приспособились к жизни в снегу или на его поверхности, выработав различные ана-томо-морфологические, физиологические, поведенческие и другие особенности. Например, у некоторых животных увеличивается к зиме опорная поверхность ног путем обрастания их жесткими волосами (рис. 5.26), перьями, роговыми щитками. т низких температур при сильных ветрах в малоснежные зимы страдают растения и животные. Так, в годы, когда снега выпадает мало, гибнут мышевидные грызуны, кроты и другие мелкие животные. Вместе с тем в широтах, где зимой выпадают осадки в виде снега, растения и животные исторически приспособились к жизни в снегу или на его поверхности, выработав различные ана-томо-морфологические, физиологические, поведенческие и другие особенности. Например, у некоторых животных увеличивается к зиме опорная поверхность ног путем обрастания их жесткими волосами (рис. 5.26), перьями, роговыми щитками. Другие мигрируют или впадают в неактивное состояние — сон, спячка, диапауза. Ряд животных переходит на питание определенными видами кормов. Б  елизна снежного покрова демаскирует темных животных. Сезонная смена окраски у белой и тундряной куропаток, горностая (рис. 5.27), зайца-беляка, ласки, песца, несомненно, связана с отбором на маскировку под цвет фона. елизна снежного покрова демаскирует темных животных. Сезонная смена окраски у белой и тундряной куропаток, горностая (рис. 5.27), зайца-беляка, ласки, песца, несомненно, связана с отбором на маскировку под цвет фона.Осадки помимо непосредственного воздействия на организмы обусловливают ту или иную влажность воздуха, которая, как уже отмечалось, играет важную роль в жизни растений и животных, так как влияет на интенсивность их водного обмена. Испарение с поверхности тела животных и транспирация у растений идут тем интенсивнее, чем меньше воздух насыщен парами воды. П Рис. 5.27. Сезонная окраска горностая оглощение надземными частями капельно-жидкой влаги, выпадающей в виде дождя, а также парообразной влаги из воздуха, у высших растений встречается у эпифитов тропических лесов, которые поглощают влагу всей поверхностью листьев и воздушных корней. Парообразную влагу из воздуха могут впитывать ветви некоторых кустарников и деревьев, например саксаулов — Halaxylon persicum, H. aphyllum. У высших споровых и особенно низших растений поглощение влаги надземными частями является обычным способом водного питания (мхи, лишайники и др.). При недостатке влаги мхи, лишайники способны переживать длительное время в состоянии, близком к воздушно-сухому, впадая в анабиоз. Но стоит пройти дождю, как эти растения быстро впитывают влагу всеми наземными частями, приобретают мягкость, восстанавливают тургор, возобновляют процессы фотосинтеза и роста. У растений сильно увлажненных наземных сред обитания нередко возникает необходимость удаления избытка влаги. Как правило, это бывает, когда почва хорошо прогрета и корни активно всасывают воду, а транспирация отсутствует (утром или при тумане, когда влажность воздуха 100%). Избыточная влага удаляется путем гуттации — это выделение воды через специальные выделительные клетки, расположенные по краю или на острие листа (рис. 5.28). 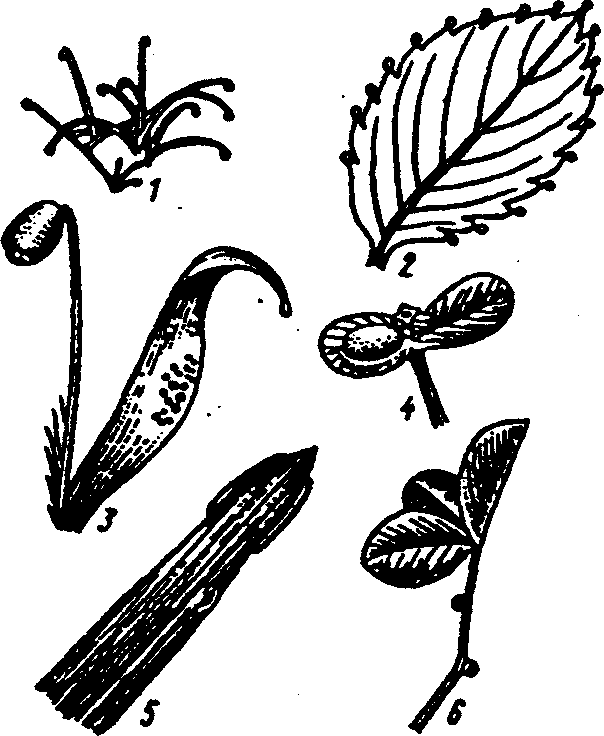 Рис. 5.28. Типы гуттации у разных растений (по А.М. Гродзинскому, 1965): 1 — у злаков, 2 — у земляники, 3 — у тюльпана, 4 — у молочая, 5 — у беллевалии сарматской, 6 — у клевера К гуттации способны не только гигрофиты, но и многие мезофиты. Например, в украинских степях гуттация обнаружена более чем у половины всех видов растений. Многие луговые травы гутгируют так сильно, что увлажняют поверхность почвы. Так животные и растения приспосабливаются к сезонному распределению осадков, к их количеству и характеру. Этим определяется состав растений и животных, сроки протекания тех или иных фаз в цикле их развития. На влажность оказывает влияние и конденсация водяных паров, часто происходящая в приземном слое воздуха при смене температуры. Выпадение росы проявляется при снижении температуры в вечерние часы. Нередко роса выпадает в таком количестве, что обильно смачивает растения, стекает в почву, увеличивает влажность воздуха и создает благоприятные условия для живых организмов, особенно когда других осадков выпадает мало. Осаждению росы способствуют растения. Охлаждаясь ночью, они конденсируют на себе водяные пары. На режим влажности значительно влияют туманы, густая облачность и другие природные явления. При количественной характеристике среды обитания растений по водному фактору используют показатели, отражающие содержание, распределение влаги не только в воздухе, но и в почве. Почвенная вода, или влажность почвы, является одним из основных источников влаги для растений. Вода в почве находится в раздробленном состоянии, вкраплена в поры разных размеров и форм, имеет большую поверхность раздела с почвой, содержит ряд катионов и анионов. Отсюда почвенная влага неоднородна по физическим и химическим свойствам. Не вся вода, содержащаяся в почве, может быть использована растениями. По физическому состоянию, подвижности, доступности и значению для растений почвенная вода подразделяется на гравитационную, гигроскопическую и капиллярную. В почве содержится и парообразная влага, занимающая все свободные от воды поры. Это почти всегда (кроме пустынных почв) насыщенный водяной пар. При понижении температуры ниже 0°С почвенная влага переходит в лед (вначале свободная вода, а при дальнейшем охлаждении — и часть связанной). Общее количество воды, которое может быть удержано почвой (его определяют, добавляя избыток воды и затем ожидая, пока она не перестанет выходить каплями), называется полевой влагоемкостью. Содержание влаги в почве, при котором растение не удовлетворяет свою потребность в воде, называется коэффициентом за-вядания. Для одйого и того же вида растения на разных почвах коэффициент завядания неодинаков и составляет, например, для тяжелой глины 16,3%, а для крупного песка— 0,9%. Следовательно, общее количество воды в почве не может характеризовать степень обеспеченности растений влагой. Для ее определения из общего количества воды необходимо вычесть коэффициент завядания. Однако физически доступная вода почвы физиологически не всегда доступна растениям из-за низкой температуры почвы, недостатка кислорода в почвенной воде и почвенном воздухе, кислотности почвы, высокой концентрации растворенных в почвенной воде минеральных солей. Несоответствие между всасыванием воды корнями и отдачей ее листьями приводит к завяданию растений. От количества физиологически доступной воды зависит развитие не только надземных частей, но и корневой системы растений. У растений, произрастающих на сухих почвах, корневая система, как правило, более разветвлена, более мощная, чем на влажных (рис. 5.29). 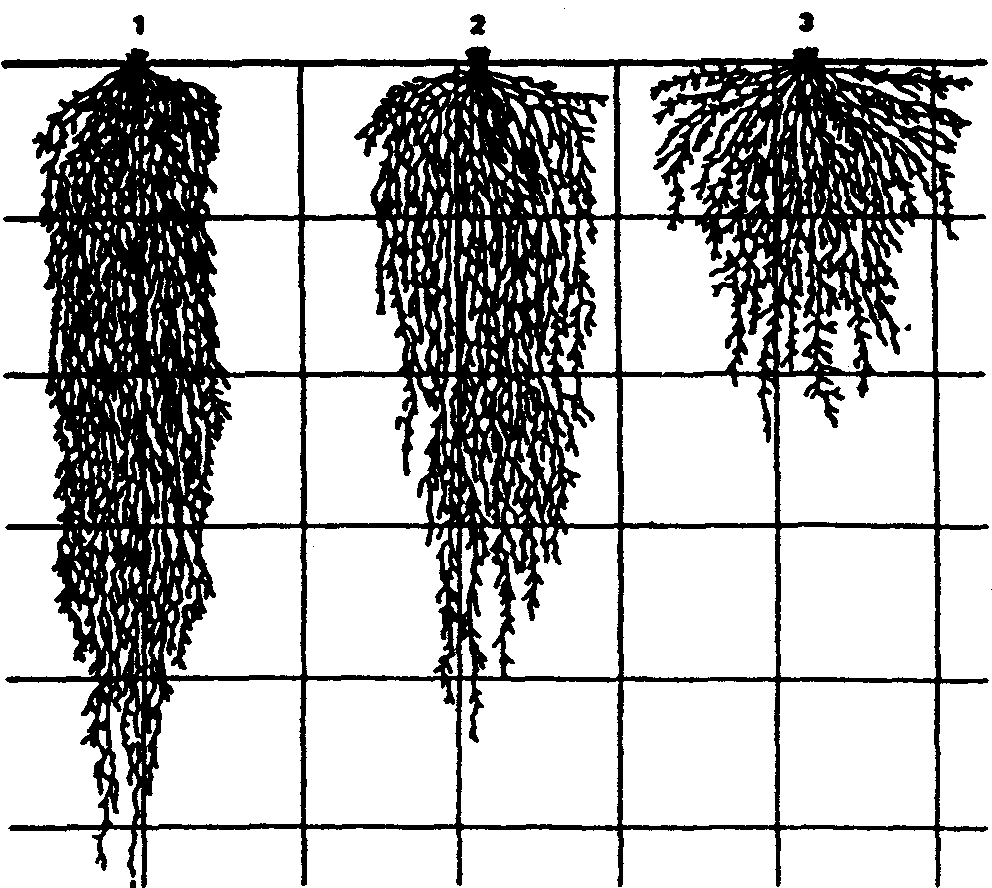 Рис. 5.29. Корневая система озимой пшеницы (по В. Г. Хржановскому и др., 1994): 1 — при большом количестве осадков; 2 — при среднем; 3 — при малом Одним из источников почвенной влаги являются грунтовые воды. При низком их уровне капиллярная вода не достигает почвы и не влияет на ее водный режим. Увлажнение почвы за счет только атмосферных осадков вызывает сильные колебания ее влажности, что часто отрицательно влияет на растения. Вредно сказывается и слишком высокий уровень грунтовых вод, потому что это приводит к переувлажнению почвы, к обеднению кислородом и обогащению минеральными солями. Постоянное увлажнение почвы независимо от капризов погоды обеспечивает оптимальный уровень грунтовых вод. Температурный режим. Отличительной чертой наземно-воздушной среды является большой размах температурных колебаний. В большинстве районов суши суточные и годовые амплитуды температур составляют десятки градусов. Особенно значительны изменения температуры воздуха в пустынях и приполярных континентальных районах. Например, сезонный размах температуры в пустынях Средней Азии 68—77°С, а суточный 25— 38°С. В окрестностях Якутска среднеянварская температура воздуха-43°С, среднеиюльская +19°С, а годовой размах от-64 до +35°С. В Зауралье годовой ход температуры воздуха резкий и сочетается с большой изменчивостью температур зимних и весенних месяцев в разные годы. Самым холодным является январь, средняя температура воздуха составляет от -16 до -19°С, в отдельные годы понижается до -50°С, самый теплый месяц июль с температурой от 17,2 до 19,5°С. Максимальные плюсовые температуры 38—41°С. Еще более значительны колебания температуры на поверхности почвы. Наземные растения занимают зону, прилежащую к поверхности почвы, т. е. к «поверхности раздела», на которой совершается переход падающих лучей из одной среды в другую или по-другому — из прозрачной в непрозрачную. На этой поверхности создается особый тепловой режим: днем — сильное нагревание благодаря поглощению тепловых лучей, ночью — сильное охлаждение вследствие лучеиспускания. Отсюда приземный слой воздуха испытывает наиболее резкие суточные колебания температур, которые в наибольшей степени выражены над оголенной почвой. Тепловой режим местообитания растений, например, характеризуется на основе измерений температуры непосредственно в растительном покрове. В травянистых сообществах измерения делают внутри и на поверхности травостоя, а в лесах, где существует определенный вертикальный градиент температуры, — в ряде точек на разных высотах. Устойчивость к температурным изменениям среды у наземных организмов различна и зависит от конкретного местообитания, где протекает их жизнь. Так, наземные листостебельные растения в большинстве своем растут в широком температурном диапазоне, т. е. являются эвритермными. Их жизненный интервал в активном состоянии простирается, как правило, от 5 до 55°С, при этом между 5 и 40°С эти растения продуктивны. Растения континентальных областей, для которых характерен четкий суточный ход температуры, развиваются лучше всего, когда ночь на 10—15°С холоднее, чем день. Это относится к большинству растений умеренной зоны — при разнице температур 5—10°С, а тропические растения при еще меньшей амплитуде — около 3°С (рис. 5.30). 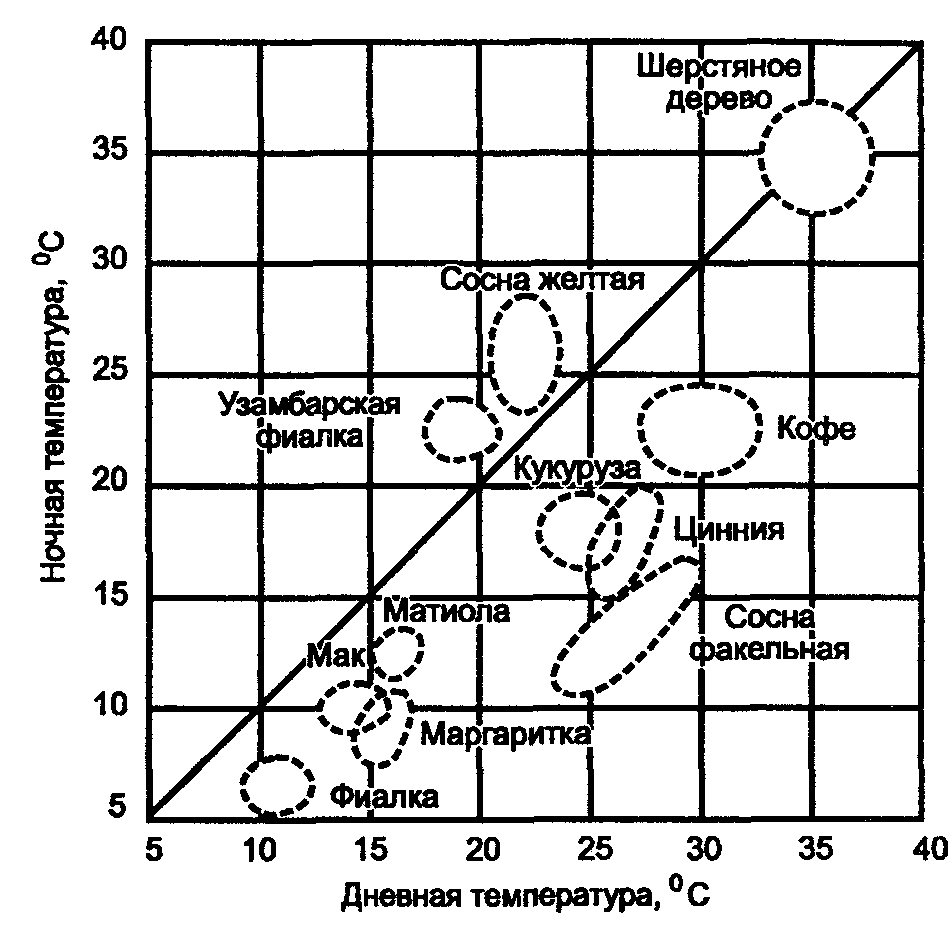 Рис. 5.30. Области оптимальных температур для роста и развития различных растений (по Went, 1957) У пойкилотермных организмов с повышением температуры (Т) продолжительность развития (t) уменьшается все быстрее. Скорость развития Vt может быть выражена формулой Vt = 100/t. Для достижения определенной стадии развития (например, у насекомых — из яйца), т.е. окукливания, имагинальной стадии, всегда требуется определенная сумма температур. Произведение эффективной температуры (температуры выше нулевого пункта развития, т. е. Т—То) на длительность развития (t) дает специфическую для данного вида термалъную постоянную развития c=t(T—То). Используя данное уравнение, можно рассчитать время наступления определенной стадии развития, например, вредителя растения, на которой эффективна с ним борьба. Растения как пойкилотермные организмы не имеют собственной стабильной температуры тела. Их температура определяется тепловым балансом, т. е. соотношением поглощения и отдачи энергии. Эти величины зависят от многих свойств как окружающей среды (размеры прихода радиации, температура окружающего воздуха и его движения), так и самих растений (окраска и другие оптические свойства растения, величина и расположение листьев и др.). Первостепенную роль играет охлаждающее действие транспирации, которая препятствует сильным перегревам растений в жарких местообитаниях. Как результат действия вышеуказанных причин, температура растений обычно отличается (нередко довольно значительно) от температуры окружающего воздуха. Здесь возможны три ситуации: температура растения выше температуры окружающего воздуха, ниже ее, равна или очень близка к ней. Превышение температуры растений над температурой воздуха встречается не только в сильно прогреваемых, но и в более холодных местообитаниях. Этому способствуют темная окраска или иные оптические свойства растений, которые увеличивают поглощение солнечной радиации, а также анатомо-морфологические особенности, способствующие снижению транспирации. Довольно заметно могут нагреваться арктические растения (рис. 5.31). Другим примером является карликовая ива — Salix arctica на Аляске, у которой днем листья теплее воздуха на 2—11 С и даже в ночные часы полярного «круглосуточного дня» — на 1—3°С. Ранневесенним эфемероидам, так называемым «подснежникам», нагревание листьев обеспечивает возможность достаточно интенсивного фотосинтеза в солнечные, но еще холодные весенние дни. Для холодных местообитаний или связанных с сезонными колебаниями температур повышение температуры растений экологически очень важно, так как физиологические процессы при этом получают независимость в известных пределах от окружающего теплового фона. 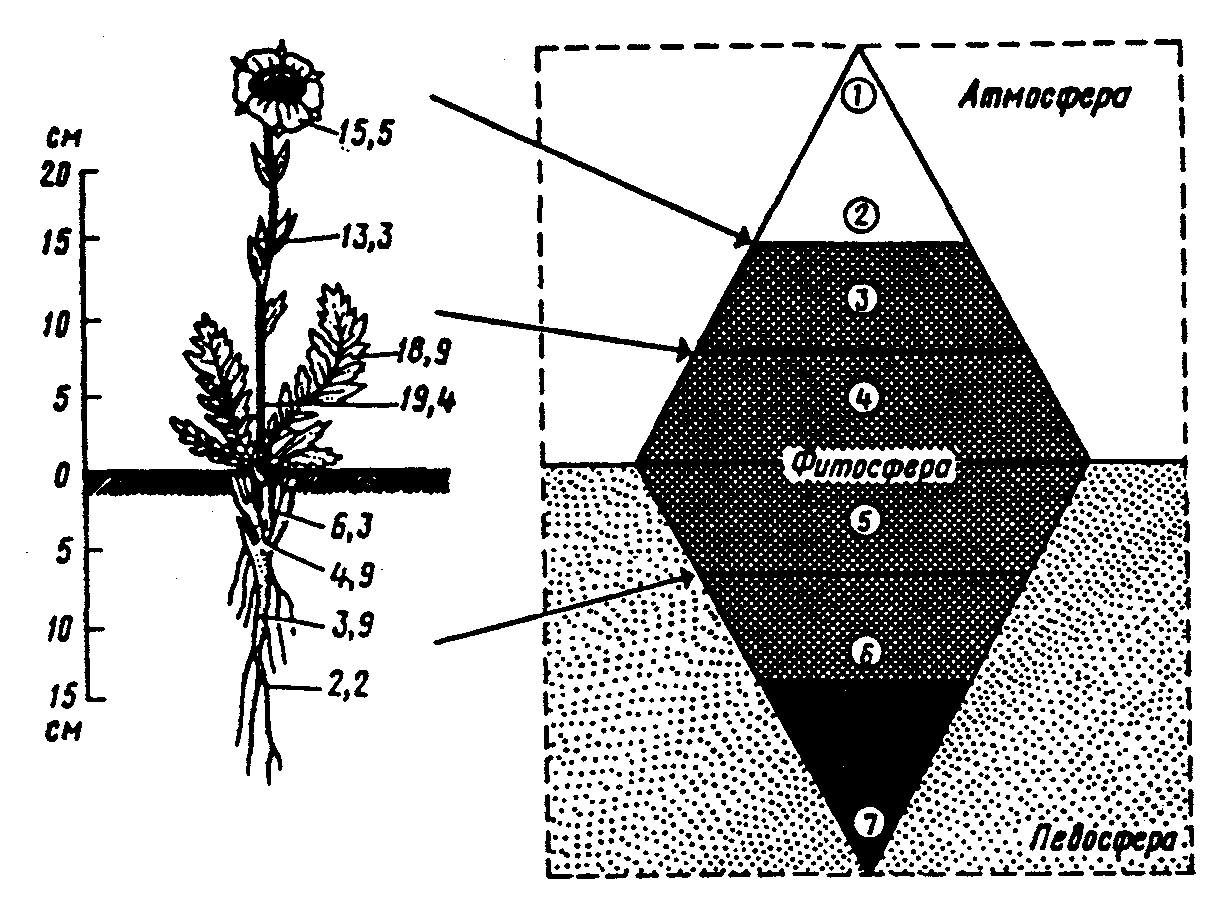 Рис. 5.31. Распределение температур в розеточном растении арктической тундры (Novosieversia glacialis) в солнечное июньское утро при температуре воздуха 11,7°С (по Б. А. Тихомирову, 1963) Справа — интенсивность процессов жизнедеятельности в биосфере: 1 — самый холодный слой воздуха; 2 — верхняя граница прироста побегов; 3, 4, 5 — зона наибольшей активности жизненных процессов и максимального накопления органического вещества; 6 — уровень вечной мерзлоты и нижняя граница укоренения; 7 — область наиболее низких температур почвы Снижение температуры растений по сравнению с окружающим воздухом чаще всего отмечается в сильно освещенных и прогреваемых участках наземной сферы (пустыня, степь), где листовая поверхность растений сильно редуцирована, а усиленная транспирация способствует удалению избытка тепла и предотвращает перегрев. В общих чертах можно сказать, что в жарких местообитаниях температура надземных частей растений ниже, а в холодных — выше температуры воздуха. Совпадение температуры растений с температурой окружающего воздуха встречается реже — в условиях, исключающих сильный приток радиации и интенсивную транспира-цию, например, у травянистых растений под пологом лесов, а на открытых участках — в пасмурную погоду или при дожде. В целом же наземные организмы по сравнению с водными отличаются большей эвритермностью. В наземно-воздушной среде условия жизни осложняются существованием погодных изменений. Погода — это непрерывно меняющееся состояние атмосферы у земной поверхности, примерно до высоты 20 км (граница тропосферы). Изменчивость погоды проявляется в постоянном варьировании сочетания таких факторов среды, как температура и влажность воздуха, облачность, осадки, сила и направление ветра и т. д. (рис. 5.32). 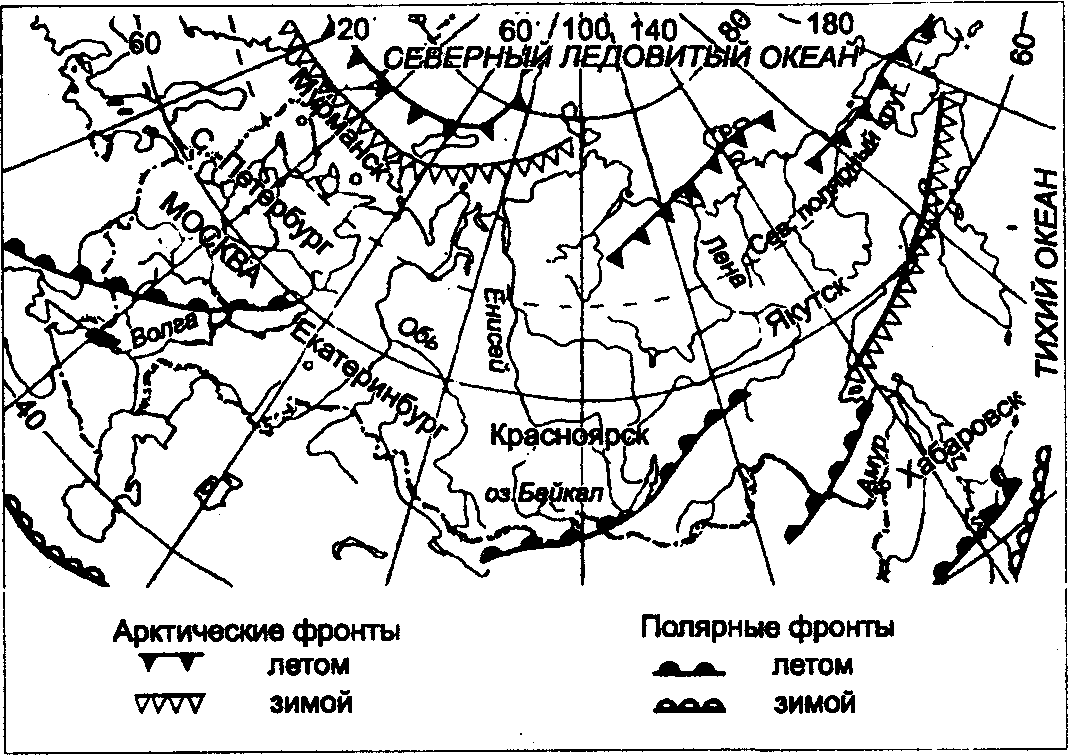 Рис. 5.32. Атмосферные фронты над территорией России Для погодных изменений наряду с закономерным чередованием их в годовом цикле характерны непериодические колебания, существенно усложняющие условия существования наземных организмов. На рис. 5.33 на примере гусеницы яблоневой плодожорки Carpocapsa pomonella показана зависимость смертности от температуры и относительной влажности. 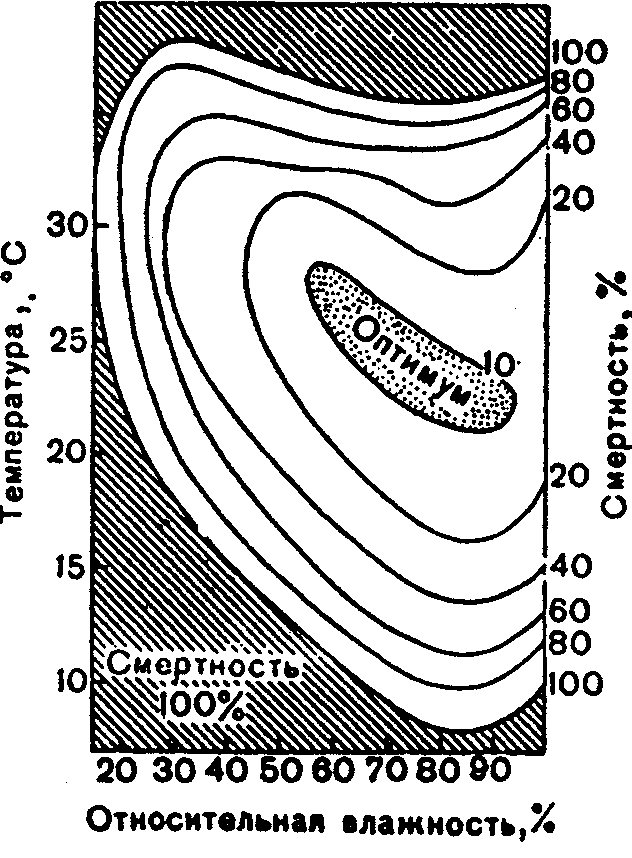 Рис. 5.33. Смертность гусениц яблоневой плодожорки Саrpocapsa pomonella в зависимости от температуры и влажности (по Р. Дажо,1975) Из нее следует, что кривые равной смертности имеют концентрическую форму и что оптимальная зона ограничена относительной влажностью 55 и 95% и температурой 21 и 28°С. Свет, температура и влажность воздуха обусловливают у растений обычно не максимальную, а среднюю степень открытия устьиц, так как совпадение всех условий, способствующих их открытию, случается редко. Многолетний режим погоды характеризует климат местности. В понятие климата входят не только средние значения метеорологических явлений, но и их годовой и суточный ходы, отклонение от него, их повторяемость. Климат определяется географическими условиями района. Основные климатические факторы — это температура и влажность, измеряемые количеством осадков и насыщенностью воздуха водяными парами. Так, в удаленных от моря странах наблюдается постепенный переход от гумидного климата через семиаридную промежуточную зону со случайными или периодическими засушливыми периодами к аридной территории, для которой характерны продолжительная засуха, засоление почвы и воды (рис. 5.34). 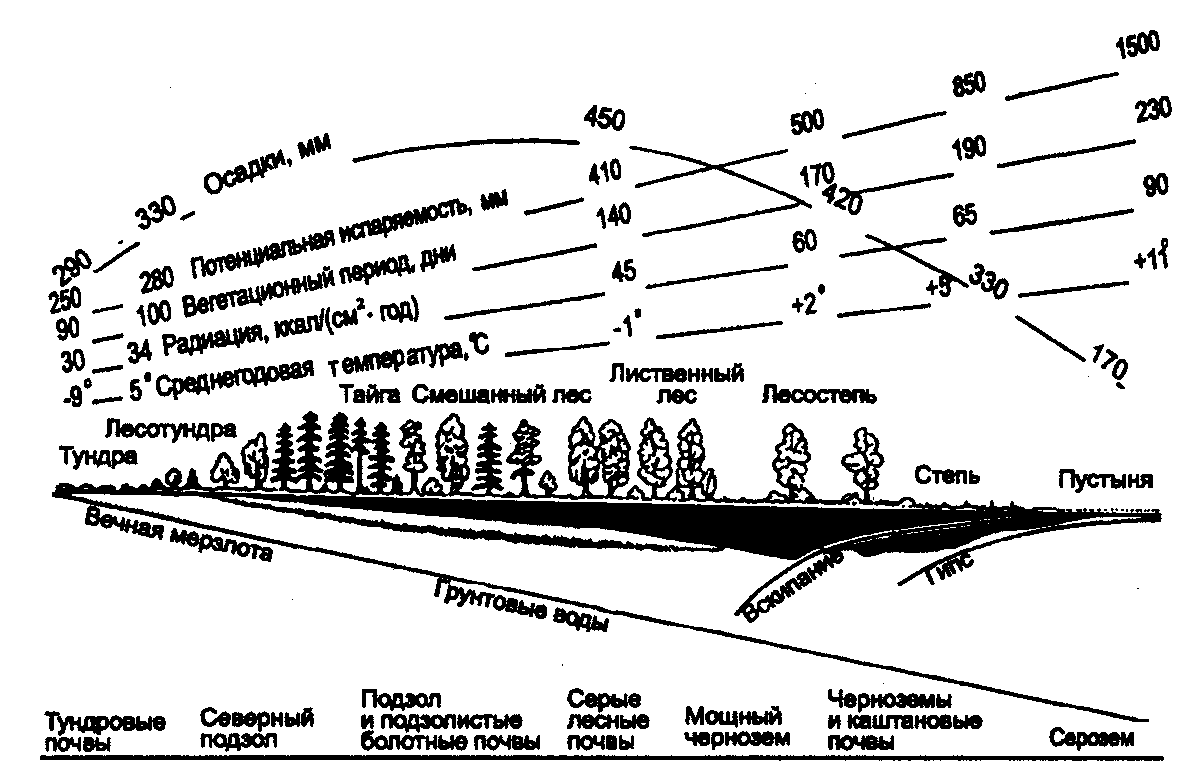 Рис. 5.34. Схема изменения климата, растительности и почв вдоль профиля через основные ландшафты Европейской части России с северо-запада на юго-восток до Прикаспийской низменности (по В. Н. Сукачеву, 1934) Примечание: там, где кривая осадков пересекает восходящую линию испаряемости, расположена граница между гумидным (слева) и аридным (справа) климатом. Черным показан гумусовый горизонт, штриховкой — иллювиальный горизонт Каждое местообитание характеризуется определенным экологическим климатом, т. е. климатом приземного слоя воздуха, или экоклиматом. Большое влияние на климатические факторы оказывает растительность. Так, под пологом леса влажность воздуха всегда выше, а колебания температуры меньше, чем на полянах. Отличается и световой режим этих мест. В разных растительных ассоциациях формируется свой режим света, температуры, влажности, т. е. своеобразный фитоклимат. Для полной характеристики климатических условий того или иного местообитания не всегда достаточно данных экоклимата или фитоклимата. Местные элементы среды (рельеф, экспозиция, растительность и т. п.) очень часто так изменяют в конкретном участке режим света, температуры, влажности, движение воздуха, что он значительно может отличаться от климатических усло-вий местности. Локальные модификации климата, складывающиеся в приземном слое воздуха, называют микроклиматом. Например, условия жизни, окружающие личинок насекомых, живущих под корой дерева, иные, чем в лесу, где это дерево растет. Температура южной стороны ствола может быть на 10 — 15°С выше температуры ее северной стороны. Устойчивым микроклиматом обладают заселенные животными норы, дупла деревьев, пещеры. Четких же различий между экоклиматом и микроклиматом не существует. Считается, что экоклимат — это климат больших территорий, а микроклимат — климат отдельных небольших участков. Микроклимат оказывает влияние на живые организмы той или иной территории, местности (рис. 5.35). 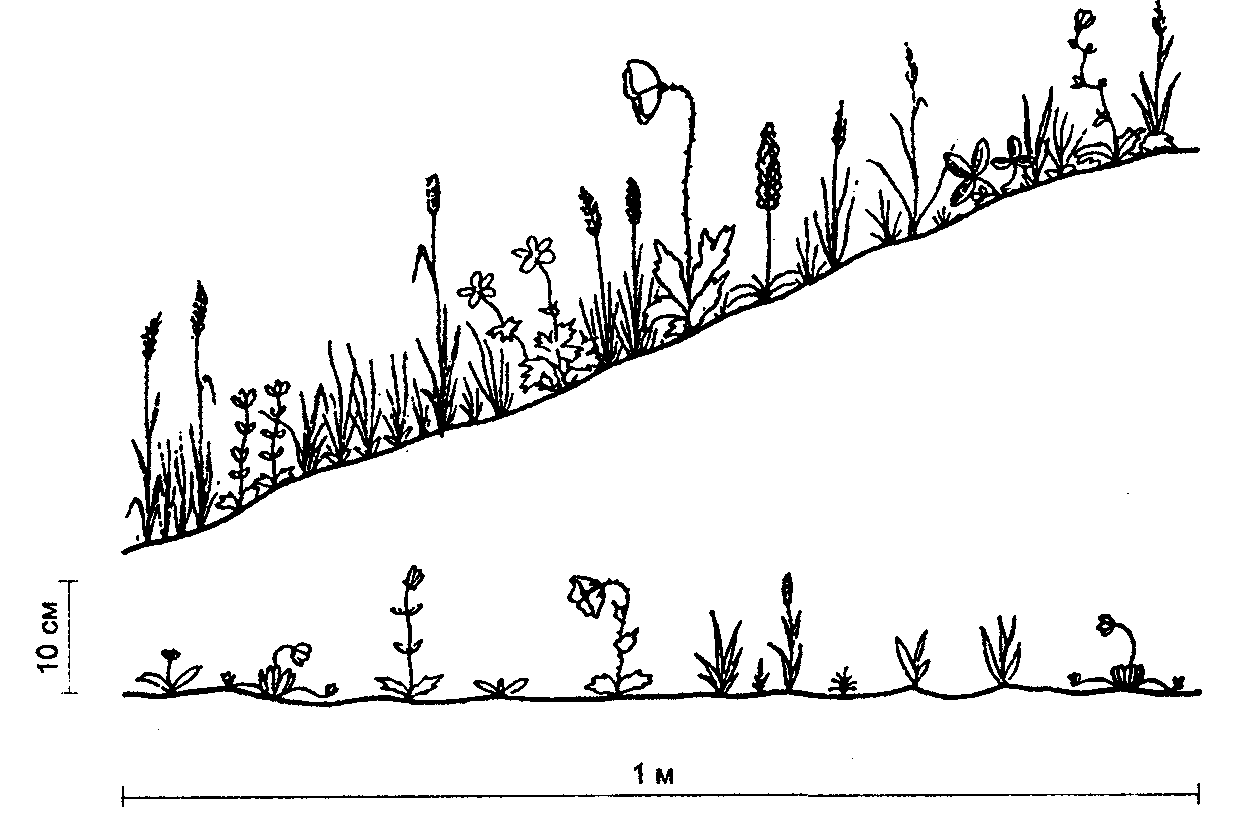 Рис. 5.35. Влияние микроклимата на растительность в тундре (по Ю. И. Чернову, 1979): вверху — хорошо прогреваемый склон южной экспозиции; внизу — горизонтальный участок плакора (флористический состав на обоих участках одинаков) Наличие в одной местности многих микроклиматов обеспечивает сосуществование видов, обладающих неодинаковыми требованиями к внешней среде. Географическая поясность и зональность. Распространение живых организмов на Земле тесно связано с географическими поясами и зонами. Пояса имеют широтное простирание, что, естественно, обусловлено в первую очередь радиационными рубежами и характером атмосферной циркуляции. На поверхности земного шара выделяют 13 географических поясов, имеющих распространение на материках и океанах (рис. 5.36). 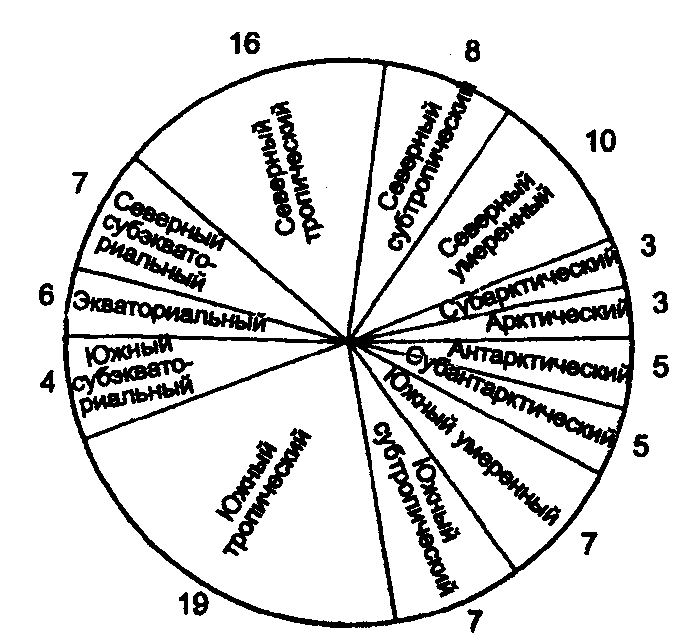 Рис. 5.36. Соотношение площадей суши, занятых различными физико-географическими поясами, в % (по Н. Ф. Реймерсу, 1990) Это такие, как арктический, антарктический, субарктический, субантарктический, северный и южный умеренные, северный и южный субарктические, северный и южный тропические, северный и южный субэкваториальные и экваториальный. Внутри поясов выделяют географические зоны, где наравне с радиационными условиями принимаются во внимание увлажнение емной поверхности и соотношение тепла и влаги, свойственные данной зоне. В отличие от океана, где обеспеченность влагой полная, на материках соотношение тепла и влаги может иметь значительные отличия. Отсюда географические пояса распространяются на материки и океаны, а географические зоны — только на материки. Различают широтные и меридиальные или долготные природные зоны. Первые тянутся с запада на восток, вторые — с севера на юг. В долготном направлении широтные зоны подразделяются на подзо-ны, а в широтном — на провинции. Основоположником учения о природной зональности является В. В. Докучаев (1846—1903), который обосновал зональность как всеобщий закон природы. Этому закону подчинены все явления в пределах биосферы. Основные причины зональности — форма Земли и ее положение относительно солнца. На распределение тепла на Земле помимо широтности влияют характер рельефа и высота местности над уровнем моря, соотношение суши и моря, морские течения и др. В дальнейшем радиационные основь^ формирования зональности земного шара были разработаны А. А. Григорьевым и М. И. Будыко. Для установления количественной характеристики соотношения тепла и влаги для различных географических зон ими были определены некоторые коэффициенты. Соотношение тепла и влаги выражено отношением радиационного баланса поверхности к скрытой теплоте испарения и сумме осадков (радиационный индекс сухости). Был установлен закон, получивший название закона периодической географической зональности (А. А. Григорьева — М. И. Будыко), который гласит, что со сменой географических поясов аналогичные географические (ландшафтные, природные) зоны и их некоторые общие свойства периодически повторяются. Каждая зона приурочена к определенному интервалу значений-показателей: особый характер геоморфологических процессов, особый тип климата, растительности, почв и животного мира. На территории бывшего СССР отмечали следующие географические зоны: ледяную, тундры, лесотундры, тайги, смешанных лесов. Русской равнины, муссонных смешанных лесов Дальнего Востока, лесостепей, степей, полупустынь, пустынь умеренного пояса, пустынь субтропического пояса, средиземноморского и влажных субтропиков. Одним из важных условий изменчивости организмов и их зонального размещения на земле служит изменчивость химического состава среды. В этом отношении большое значение имеет учение А. П. Виноградова о биогеохимических провинциях, которые определяются зональностью химического состава почв, а также климатической, фитогеографической и геохимической зональностью биосферы. Биогеохимические провинции — это области на поверхности Земли, различающиеся по содержанию (в почвах, водах и т. д.) химических соединений, с которыми связаны определенные биологические реакции со стороны местной флоры и фауны. Наряду с горизонтальной зональностью в наземной среде четко проявляется высотная или вертикальная поясность. Растительность горных стран более богата, чем на прилегающих равнинах, и характеризуется повышенным распространением эндемических форм. Так, по данным О. Е. Агаханянца (1986), флора Кавказа насчитывает 6350 видов, из которых 25% эндемич-ны. Флора гор Средней Азии оценивается в 5500 видов, из них 25—30% эндемики, в то время как на прилегающих равнинах юж-ных пустынь насчитывается 200 видов растений. При подъеме в горы повторяется та же смена зон, что и от экватора к полюсам. У подножия обычно располагаются пустыни, затем степи, широколиственные леса, хвойные леса, тундра и, наконец, льды. Однако полной аналогии все же нет. При подъеме в горы понижается температура воздуха (средний градиент температуры воздуха 0,6 °С на 100 м), снижается испаряемость, усиливаются ультрафиолетовая радиация, освещенность и т. д. Все это заставляет растения приспосабливаться к сухой или влажной вреде. Здесь доминируют среди растений подушкообразные жизненные формы, многолетники, у которых выработана адаптация к сильной ультрафиолетовой радиации и снижению транспирации. Своеобразен и животный мир высокогорных районов. Пониженное давление воздуха, значительная солнечная радиация, резкие колебания дневных и ночных температур, изменение влажности воздуха с высотой способствовали выработке специфических физиологических адаптации организма горных животных. Например, у животных увеличивается относительный объем сердца, возрастает содержание гемоглобина в крови, что позволяет более интенсивно поглощать кислород из воздуха. Каменистый грунт осложняет или почти исключает норовую деятельность животных. Многие мелкие животные (мелкие грызуны, пищухи, ящерицы и др.) находят убежища в расщелинах скал, в пещерах. Из птиц для горных районов характерны горные индейки (улары), горные вьюрки, жаворонки, из крупных птиц — бородачи, грифы, кондоры. В горах из крупных млекопитающих обитают бараны, козлы (в том числе и снежные козлы), серны, яки и др. Хищники представлены такими видами, как волки, лисицы, медведи, рыси, снежный барс (ирбис) и т. д. |