эндокринная система. Эндокринная система
 Скачать 55.04 Kb. Скачать 55.04 Kb.
|

|
Константинова Анна 14 группа 1 курс СФ Эндокринная система Контроль гипофиза гипоталамусом Гипофиз (питуитарная железа, нижний мозговой придаток) — железа внутренней секреции, расположенная у основания головного мозга. Этот орган размером с горошину можно назвать «мозговым центром» всей эндокринной системы нашего организма. Гипофиз, будучи эндокринным органом, обладает разнообразными функциями, которые осуществляются при помощи гормонов его передней и задней долей, а также промежуточной части. Ряд гормонов передней доли называется тройными (напр., тиреотропный гормон). В передней доле гипофиза вырабатываются гормоны: -тиреотропный гормон , -адренокортикотропный гормон, -гормон роста (Соматотропный гормон), -Пролактин , -фолликулостимулирующий гормон, -лютеинизирующий гормон, -липотропные факторы гипофиза. В промежуточной части образуется меланоцитостимулирующий гормон, а в задней доле накапливается вазопрессин и окситоцин. Тесно связанный через гипоталамус со всей нервной системой, гипофиз объединяет в функциональное целое эндокринную систему, участвующую в обеспечении постоянства внутренней среды организма. Понятие «постоянство» включает не только процесс поддержания основных констант внутренней среды, но и наиболее адекватное, оптимальное вегетативное обеспечение биологических функций организма, постоянное обеспечение готовности к действию. Поскольку изменяющиеся условия окружающей среды диктуют необходимость различных по биологическому значению и моторным проявлениям поведенческих реакций, то и параметры внутренней среды должны также адекватно изменяться. Известны суточные (циркадные), месячные, сезонные и другие биоритмические колебания параметров внутренней среды, в частности концентраций гормонов. Можно говорить о гомеостатическом поддержании постоянства гормонов в крови и о гомеокинетических механизмах изменений их концентрации. Внутри эндокринной системы гомеостатическая регуляция осуществляется на основе универсального принципа отрицательной обратной связи. Факт существования такой связи между передней долей гипофиза и «железами-мишенями» (щитовидная железа, кора надпочечников, гонады) твердо установлен многочисленными исследованиями. Избыток гормона «железы-мишени» тормозит, а его недостаток стимулирует секрецию и выделение соответствующего тронного гормона. В петлю обратной связи непременно включается гипоталамус: именно в нем находятся чувствительные к концентрации в крови гормонов «желез-мишеней» рецепторные зоны. Улавливая отклонения концентраций гормонов от необходимого уровня, рецепторы гипоталамуса активизируют или тормозят соответствующие Гипоталамические центры, управляющие работой передней доли гипофиза путем выделения соответствующих гипоталамических аденогипофизарных гормонов. Увеличивая или снижая продукцию тропных гормонов, гипофиз устраняет отклонения функции «железы-мишени». Основное свойство регуляции по отклонению состоит в том, что сам факт отклонения концентрации гормонов «желез-мишеней» от нормы является стимулом для возврата этих концентраций к заданному уровню. В свою очередь «заданный уровень» не является постоянной величиной в течение длительного времени. Он изменяется, порой значительно, за счет гомеокинетических механизмов, переводящих его на новый заданный уровень, в дальнейшем столь же строго поддерживаемый регуляцией «по отклонению». Гомеокинетической перестройкой можно объяснить сезонные изменения концентрации гормонов в крови, овариально-менструальный цикл, циркадные колебания количества оксикетостероидов и т. д. В основе гомеокинеза лежит регуляция «по возмущению». Не имеющий прямого отношения к концентрации гормона, возмущающий фактор (температура окружающей среды, длительность светового дня, стрессовая ситуация и т. п.) воздействует на ЦНС, через органы чувств, в т. ч. и на те ядра гипоталамуса, которые управляют работой передней доли гипофиза. Именно в них и происходит «перестройка уровня», адекватно соответствующая будущей деятельности. В процессе гомеостатической регуляции «по отклонению» и в процессе гомеокинетической регуляции «по возмущению» гипоталамогипофизарный комплекс выступает как единое, неразрывное целое. Поскольку Гипофиз является важнейшим звеном в системе соматовегетативной интеграции, нарушения его функции ведут к дискоординациям вегетативной и соматической сферы. Регуляция секреции альдостерона. Регуляция секреции альдостерона так тесно переплетается с регуляцией концентрации электролитов во внеклеточной жидкости, объема внеклеточной жидкости, объема крови, артериального давления и многих специфических функций почек, что трудно обсуждать регуляцию продукции альдостерона независимо от этих факторов. Регуляция секреции альдостерона клубочковой зоной почки независит от регуляции продукции кортизола и андрогенов пучковой и сетчатой зон. Известны четыре фактора, играющие наиболее важную роль в продукции альдостерона. 1. Увеличение концентрации калия во внеклеточной жидкости резко повышает продукцию альдостерона. 2. Увеличение активности ренин-ангиотензиновой системы (увеличение уровня ангиотензина II) также существенно повышает секрецию альдостерона. 3. Увеличение концентрации натрия во внеклеточной жидкости незначительно снижает продукцию альдостерона. 4. АКТГ аденогипофиза необходим для синтеза альдостерона, но оказывает слабое влияние на регуляцию скорости его секреции. Из всех этих факторов концентрация ионов калия и ренин-ангиотензиновая система наиболее активны в регуляции продукции альдостерона. Увеличение концентрации калия всего на несколько процентов увеличивает продукцию альдостерона во много раз. Сходным образом активация ренин-ангиотензиновой системы обычно в ответ на снижение объемного кровотока в почке или потерю натрия могут вызвать многократное увеличение продукции альдостерона. В свою очередь, альдостерон действует на почки: (1) помогает им экскретировать избыток ионов калия; (2) увеличивает объем крови и артериальное давление, возвращая активность ренин-ангиотензиновой системы к нормальному уровню. В данном случае механизм обратной связи жизненно важен. На рисунке ниже показано влияние блокады образования ангиотензина II, вызванной ферментом, ингибирующим превращение ангиотензина на фоне пребывания человека на гипонатриевой диете в течение нескольких недель, которая обычно приводит к увеличению концентрации альдостерона в несколько раз. 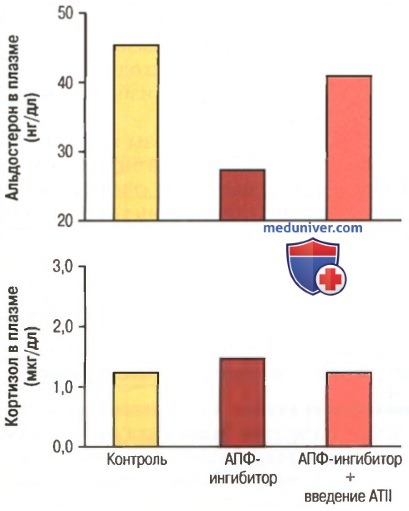 Заметим, что блокада выработки ангиотензина II существенно снижает концентрацию альдостерона без заметных изменений концентрации кортизола; это указывает на важную роль ангиотензина II в стимуляции секреции альдостерона, когда снижены поступления натрия и объем внеклеточной жидкости. Напротив, сами влияния концентрации ионов натрия и АКТГ на продукцию альдостерона обычно минимальны. Тем не менее, снижение внеклеточной концентрации ионов натрия на 10-20%, что случается чрезвычайно редко, может удвоить продукцию альдостерона. Даже малого количества АКТГ, продуцируемого аденогипофизом, достаточно для секреции необходимого количества альдостерона, но полное отсутствие АКТГ может значительно снизить синтез альдостерона клубочковой зоной. 3) Регуляция секреции гормонов щитовидной железы Для обеспечения нормального уровня метаболической активности необходима постоянная продукция гормонов, соответствующая потребностям организма; это соответствие обеспечивается специальным механизмом обратной связи, опосредованным гипоталамусом и передней долей гипофиза, контролирующим уровень секреции, в частности щитовидных желез. Основные этапы регуляции следующие. Тиреотропный гормон (гормон аденогипофиза) стимулирует секрецию щитовидных желез. ТТГ, известный также как тиреотропин, является гормоном передней доли гипофиза (гликопротеином с молекулярной массой около 28000). Этот гормон увеличивает продукцию тироксина и трийодтиронина щитовидными железами. Его специфические влияния на щитовидные железы следующие. 1. Стимуляция протеолиза тиреоглобулинса, хранящегося в фолликулах, приводящая к высвобождению в кровоток тиреоидных гормонов и уменьшению содержимого фолликулов. 2. Стимуляция активности йодного насоса, увеличивающего скорость йодного транспорта в гландулярные клетки, а также иногда увеличивающая отношение концентрации внутриклеточного йода к концентрации внеклеточного йода более чем в 8 раз по сравнению с нормой. 3. Стимуляция йодизации тирозина и, следовательно, образования тиреоидных гормонов. 4. Увеличение размеров и стимуляция секреторной активности клеток щитовидной железы. 5. Увеличение количества тиреоидных клеток и изменение их формы с кубовидной на цилиндрическую, существенное увеличение складчатости внутренней поверхности фолликулов. В итоге ТТГ увеличивает все известные проявления секреторной активности в секреторных клетках щитовидной железы. Наиболее ранней реакцией на введение ТТГ можно считать явление протеолиза тиреоглобулина, в результате которого через 30 мин в кровоток высвобождаются тироксин и трийодтиронин. Прочие реакции требуют нескольких часов, дней и даже недель для их полной реализации. Стимулирующий эффект ТТГ опосредован аденозинмонофосфатом. Раньше было трудно объяснить многочисленные и разнообразные влияния ТТГ на тиреоидные клетки. Теперь многие, если не все эти эффекты связывают с активацией системы вторичных посредников, в данном случае представленной циклическим аденозинмонофосфатом. Первым звеном активации этой системы является связывание ТТГ со специфическим рецептором на поверхности базальной мембраны тиреоидной клетки. Это взаимодействие активирует аденилатциклазу в мембране, которая увеличивает образование цАМФ в клетке. В итоге цАМФ действует как вторичный посредник, активируя протеинкиназу, которая инициирует многочисленные процессы фосфорилирования в клетке. В результате немедленно активируются как процессы секреции, так и продолжительный рост ткани самой железы. Итак, способ регуляции активности клеток ткани щитовидной железы такой же, что и во многих других тканях-мишенях, где вторичным посредником в клетках служит цАМФ. Продукция ТТГ передней долей гипофиза регулируется тиреотропин-рилизинг гормоном гипоталамуса. Секреция ТТГ передней долей гипофиза регулируется гипоталамическим гормоном — тиреотропин-рилизинг гормоном, который высвобождается нервными окончаниями срединного возвышения гипоталамуса. Затем ТТГ из срединного возвышения транспортируется системой гипоталамо-гипофизарных портальных сосудов. ТРГ выделен в химически чистой форме. Это простое вещество — трипептидный амид (пироглутамилгистидилпролинамид). ТРГ непосредственно действует на аденогипофиз, увеличивая выход ТТГ. Если блокируется портальная система, скорость секреции ТТГ передней долей гипофиза резко снижается, но не падает до нуля. Молекулярный механизм, с помощью которого ТРГ обусловливает секрецию ТТГ, первоначально связан взаимодействием ТРГ с рецепторами на мембране клеток аденогипофиза. Это взаимодействие приводит к активации вторичных посредников, таких как ионы кальция и диацилглицерол, которые запускают каскад реакций, активирующих фосфолипазную систему вторичных посредников в клетках аденогипофиза, и приводят к продукции большого количества фосфолипазы С, что в итоге способствует выделению ТТГ. Действие холода и других нейрогенных стимулов на секрецию ТРГ и ТТГ. Одним из наиболее изученных факторов, увеличивающих продукцию ТРГ гипоталамусом и, следовательно, секрецию ТРГ передней долей гипофиза, является холодовая экспозиция. Этот эффект, несомненно, является результатом возбуждения терморегуляторных центров гипоталамуса. Помещение крыс на несколько недель в условия чрезвычайно низких температур увеличивает выброс тиреоидных гормонов, иногда более чем на 100% относительно нормы, что приводит к возрастанию основного обмена более чем на 50%. Известно, что у людей, оказавшихся в арктических регионах, основной обмен превышает нормальные показатели на 15-20%. Различные эмоциональные реакции также могут увеличить выброс ТТГ и ТРГ и опосредованно привести к увеличению продукции тиреоидных гормонов. Общее возбуждение и тревожность (факторы, резко стимулирующие симпатический отдел автономной нервной системы) могут вызвать гиперпродукцию ТТГ и, возможно, поэтому способны сместить уровень обменных процессов и повысить теплопродукцию, провоцируя извращенные реакции терморегуляторных центров. Ни эмоциональные факторы, ни низкие температуры не являются эффективными на фоне перерезки гипофизарного стебля, что указывает на опосредованность этих реакций гипоталамусом. Роль тиреоидных гормонов и механизма обратной связи в снижении секреции ТТГ аденогипофизом. Увеличение содержания гормонов щитовидной железы в крови снижает продукцию ТТГ передней долей гипофиза. Если уровень тиреоидных гормонов превышает норму в 1,75 раза, продукция ТТГ падает практически до нуля. Почти все регуляторные проявления механизма обратной связи сохраняются, даже если аденогипофиз отделить от гипоталамуса. Вероятно, ингибирование продукции ТТГ возросшим уровнем тиреоидных гормонов осуществляется непосредственными их влияниями на переднюю долю гипофиза (для облегчения понимания просим вас изучить рисунок выше). Благодаря механизму обратной связи это влияние поддерживает в крови концентрацию свободных тиреоидных гормонов почти на постоянном уровне. |