коллоквиум нормальная физиология. коллок нф 1. Физиология, ее предмет, роль задачи и формирование врачебной деятельности. Связь физиологии с другими науками
 Скачать 1 Mb. Скачать 1 Mb.
|

|
К первым относятся рецепторы обоняния, тактильные и проприорецепторы. Они различаются тем, что преобразование энергии раздражения в энергию нервного импульса происходит у них в первом нейроне сенсорной системы. К вторично-чувствующим относятся рецепторы вкуса, зрения, слуха, вестибулярного аппарата. У них между раздражителем и первым нейроном находится специализированная рецепторная клетка, не генерирующая импульсы. Таким образом, первый нейрон возбуждается не непосредственно, а через рецепторную (не нервную) клетку. Общие механизмы возбуждения рецепторов. Рецепторы представляют собой клетки, различающие естественные раздражители и посылающие информацию о них в ЦНС. Стимуляция рецептора вызывает в дендритах изменение потенциала покоя в сторону деполяризации. При действии стимула на рецепторную клетку происходит преобразование энергии внешнего раздражения в рецепторный сигнал, или трансдукция сенсорного сигнала. Этот процесс включает в себя три основных этапа: 1) взаимодействие стимула, т. е. молекулы пахучего или вкусового вещества (обоняние, вкус), кванта света (зрение) или механической силы (слух, осязание) с рецепторной белковой молекулой, которая находится в составе клеточной мембраны рецепторной клетки; 2) внутриклеточные процессы усиления и передачи сенсорного стимула в пределах рецепторной клетки; 3) открывание находящихся в мембране рецептора ионных каналов, через которые начинает течь ионный ток, что, как правило, приводит к деполяризации клеточной мембраны рецепторной клетки (возникновению так называемого рецепторного потенциала). В первично-чувствующих рецепторах этот потенциал действует на наиболее чувствительные участки мембраны, способные генерировать потенциалы действия — электрические нервные импульсы. Во вторично-чувствующих рецепторах рецепторный потенциал вызывает выделение квантов медиатора из пресинаптического окончания рецепторной клетки. Медиатор (например, ацетилхолин), воздействуя на постсинаптическую мембрану первого нейрона, изменяет ее поляризацию (генерируется постсинаптический потенциал). Постсинаптический потенциал первого нейрона сенсорной системы называют генераторным потенциалом, так как он вызывает генерацию импульсного ответа. В первично-чувствующих рецепторах рецепторный и генераторный потенциалы — одно и то же. Генерация возбуждения в рецепторах. Возникновение рецепторного потенциала обусловлено повышением Na+-проводимости дендритов. Возникающее в них возбуждение электротонически распространяется к соме – происходит преобразование или первичная трансформация стимула в рецепторный потенциал. Поэтому рецептор представляет собой преобразователь, датчик. Возбуждение в форме рецепторного потенциала охватывает только сому. В аксоне же первичных рецепторов, начиная от аксонного холмика – места отхода аксона от сомы – происходит трансформация этого возбуждения в серию потенциалов действия. Очень важно, что после первого потенциала действия мембрана аксона гиперполяризуется существенно ниже уровня потенциала покоя. Благодаря этому обстоятельству Na+-каналы после инактивации восстанавливаются настолько, что фаза деполяризации, наступающая после первого следового гиперполяризационного потенциала, вновь достигает порогового значения, достаточного для генерации следующего потенциала действия. Следовательно, гиперполяризационный следовой потенциал служит основой формирования ритмического возбуждения нервного волокна. Во вторичных рецепторах возникает только рецепторный потенциал, а серия потенциалов действия формируется в терминалях афферентной нервной клетки, образующей контакт с рецептором. В частности, зрительные и слуховые рецепторы являются вторичными. Когда генераторный потенциал (ГП) достигает критической величины, он вызывает разряд афферентных импульсов в ближайшем перехвате Ранвье. Частота разряда прямо пропорциональна величине ГП (логарифмическая зависимость, соответствующая закону Вебера-Фехнера). Ощущение тоже увеличивается пропорционально логарифму силы раздражения. Новокаин прерывает поток этих импульсов, с чем связан его аналгетический эффект. Адаптация рецепторов - общее свойство всех рецепторов, заключающееся в приспособлении к силе раздражителя. Она проявляется в снижении чувствительности к постоянно действующему раздражителю. Человек "привыкает" к действию постоянных раздражителей - запаху, давлению одежды, звуку часов и т.п. и перестает замечать их. При адаптации снижается величина генераторного потенциала и частота импульсов, проходящих по афферентному нерву. Есть медленно адаптирующиеся рецепторы (болевые) и быстро адаптирующиеся (глаз). Не адаптируются (или почти не адаптируются) только вестибуло- и проприорецепторы. Когда действиепостоянного раздражителя прекращается, адаптация исчезает и чувствительность рецептора повышается (эффект возбуждения после торможения). Когда начинается действие какого-либо стимула рецептор реагирует на него очень энергично. По мере продолжения стимуляции рецептор адаптируется к нему, и активность в сенсорном волокне снижается до более низкого уровня. При коротких и периодических предъявлениях стимула рецептор каждый раз реагирует на него полностью, без адаптации. Медленно адаптирующиеся рецепторы служат для контроля за длительно сохраняющимися стимулами, например, степенью растяжения мышц, концентрацией Н+. Быстро адаптирующиеся рецепторы свойственны сенсорным системам, регистрация стимулов в которых происходит с высокой чувствительностью и высоким временным разрешением. Различение сигналов. Важная характеристика сенсорной системы — способность замечать различия в свойствах одновременно или последовательно действующих раздражителей. Различение начинается в рецепторах, но в этом процессе участвуют нейроны всей сенсорной системы. Оно характеризует то минимальное различие между стимулами, которое сенсорная система может заметить (дифференциальный, или разностный, порог). Зависимость силы ощущения от силы раздражения (закон Вебера—Фехнера) выражается формулой: E=a∙logI +b, где Е — величина ощущения, I — сила раздражения, а и b — константы, различные для разных модальностей стимулов. Согласно этой формуле, ощущение увеличивается пропорционально логарифму интенсивности раздражения. Абсолютную чувствительность сенсорной системы измеряют порогом реакции. Чувствительность и порог — обратные понятия: чем выше порог, тем ниже чувствительность, и наоборот. Обычно принимают за пороговую такую силу стимула, вероятность восприятия которого равна 0,5 или 0,75 (правильный ответ о наличии стимула в половине или в 3/4 случаев его действия). Более низкие значения интенсивности считаются подпороговыми, а более высокие — надпороговыми. Оказалось, что и в подпороговом диапазоне реакция на сверхслабые раздражители возможна, но она неосознаваема (не доходит до порога ощущения). Так, если снизить интенсивность вспышки света настолько, что человек уже не может сказать, видел он ее или нет, от его руки можно зарегистрировать неощущаемую кожно-гальваническую реакцию на данный сигнал. Чувствительность рецепторных элементов к адекватным раздражителям, к восприятию которых они эволюционно приспособлены, предельно высока. Так, обонятельный рецептор может возбудиться при действии одиночной молекулы пахучего вещества, фоторецептор — одиночным квантом света. Чувствительность слуховых рецепторов также предельна: если бы она была выше, мы слышали бы постоянный шум из-за теплового движения молекул. Выше упоминалось о различении силы раздражителей. Пространственное различение основано на распределении возбуждения в слое рецепторов и в нейронных слоях. Так, если два раздражителя возбудили два соседних рецептора, то различение этих раздражителей невозможно и они будут восприняты как единое целое. Необходимо, чтобы между двумя возбужденными рецепторами находился хотя бы один невозбужденный. Для временного различения двух раздражений необходимо, чтобы вызванные ими нервные процессы не сливались во времени и чтобы сигнал, вызванный вторым стимулом, не попадал в рефрактерный период от предыдущего раздражения. Амплитуда (интенсивность) стимула кодируется в виде частоты импульсов или потенциалов действия, направляющихся от рецептора в ЦНС. Повышение амплитуды стимула при условии ее надпорогового значения соответственно повышает частоту потенциалов действия. Понятие о рецептивном поле и рефлексогенных зонах Определенное множество рецепторов, связанных с отдельным афферентным волокном, называется рецептивным полем. Например, тактильное или ноцицептивное рецептивное поле кожной поверхности представляет собой разветвление одиночного чувствительного волокна. При одновременном раздражении нескольких рецепторов одного рецептивного поля с афферентного волокна можно получить лишь один ответ (от рецептора с наибольшей частотой). Возбуждение лишь одного из рецепторов может привести к возбуждению всего волокна. Однако, если на этот рецептор наносить повторное раздражение с определенным интервалом, то ответ, регистрируемый с афферентного волокна, постепенно уменьшается и исчезает. Ответ возобновится, если раздражающий стимул переместить в другую точку поля. Область расположения рецепторов, раздражение которых вызывает определенный рефлекс (например, раздражение слизистой оболочки носа - чихание) называют рефлексогенной зоной. 26) Строение и классификация синапсов. Химический синапс. Экзоцитоз медиатора в синаптическую щель и синаптический цикл. Взаимодействие медиатора с рецепторами постсинаптической и пресинаптической мембраны. Ионатропные и метаботропные рецепторы. Ионные механизмы формирования ВПСП и ТПСП. Химические синапсы. Большинство синапсов нервной системы — химические. Функционирование таких синапсов зависит от высвобождения медиаторов. Классический химический синапс представлен пресинаптической мембраной, синаптической щелью и постсинаптической мембраной. Пресинаптическая мембрана — часть булавовидного расширения нервного окончания клетки, передающей сигнал, а постсинаптическая мембрана — часть клетки, получающей сигнал. Медиатор высвобождается из булавовидного расширения посредством экзоцитоза, проходит через синаптическую щель и связывается с рецепторами на постсинаптической мембране. Под постсинаптической мембраной расположена субсинаптическая активная зона, в которой после активации рецепторов постсинаптической мембраны происходят разнообразные биохимические процессы. В булавовидном расширении расположены содержащие медиаторы синаптические пузырьки, а также большое количество митохондрий и цистерны гладкой эндоплазматической сети. Применение традиционных методик фиксации при исследовании клеток позволяет различить на пресинаптической мембране пресинаптические уплотнения, ограничивающие активные зоны синапса, к которым при помощи микротрубочек направляются синаптические пузырьки. 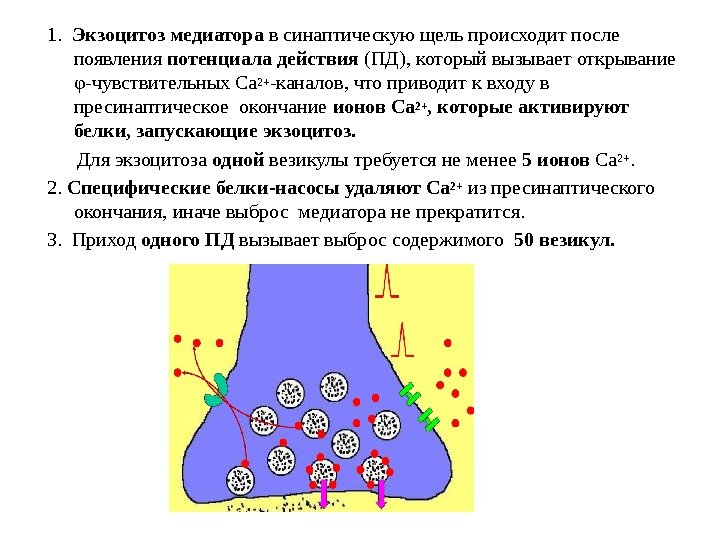 Взаимодействие медиатора с рецепторами постсинаптической мембраны. • Перемещение ионов через каналы в мембране. Это приводит к изменению заряда на ней и, следовательно, мембранного потенциала. Однонаправленность передачи - нервные сигнал передаются только от пресинаптической мембраны к постсинаптической мембране. Таким образом, синапс работает по принципу клапана, что обеспечивает надежность работы нервной системы.  В возбуждающих синапсах нервной системы медиатором может являться ацетилхолин, норадреналин, дофамин, серотонин, глугаминовая кислота, вещество Р, а также большая группа других веществ, являющихся, если не медиаторами в прямом значении, то во всяком случае модуляторами (меняющими эффектиьность) синаптической передачи. Возбуждающие медиаторы вызывают появление на постсинаптической мембраневозбуждающего постсинаптического потенциала (ВПСП). Его формирование обусловлено тем, что медиатор-рецепторный комплекс активирует Na- каналы мембраны (а также вероятно и Са-каналы) и вызывает за счет поступления натрия внутрь клетки деполяризацию мембраны. Одновременно происходит и уменьшение выхода из клетки ионовК+Амплитуда одиночного ВПСП однако довольно мала, и для уменьшения заряда мембраны до критического уровня деполяризации необходима одновременная активация нескольких возбуждающих синапсов. ВПСП, образуемые на постсинаптической мембране этих синапсов, способны суммиро-ваться, т.е. усиливать друг друга, приводя к росту амплитуды ВПСП (пространственная суммация). Растет амплитуда ВПСП и при увеличении частоты поступающих к синапсу нервных импульсов (временная суммация), что повышает число выводимых в синаптическую щель квантов медиатора. Тормозной постсинаптический потенциал (ТПСП) В тормозных синапсах обычно действуют другие, тормозные, медиаторы. Среди них хорошо изученными являются аминокислота глицин (тормозные синапсы спинного мозга), гамма-аминомасляная кислота (ГАМК) — тормозной медиатор в нейронах головного мозга. Вместе с тем, тормозной синапс может иметь тот же медиатор, что и возбуждающий, но иную природу рецепторов постсинаптической мембраны. Так, для ацетилхолина, биогенных аминов и аминокислот на постсинаптической мембране разных синапсов могут существовать как минимум два типа рецепторов, и, следовательно, разные медиатор-рецепторные комплексы способны вызывать различную реакцию хемочувствительныхрецепторуправляемых каналов. Для тор-мозного эффекта такой реакцией может являться активация калиевых каналов, что вызывает увеличение выхода ионов калия наружу и гиперполяризацию мембраны. Аналогичный эффект во многих тормозных синапсах имеет активация каналов для хлора, увеличивающая его транспорт внутрь клетки. Возникающий при гиперполяризации сдвиг мембранного потенциала получил название тормозного постсинаптического. 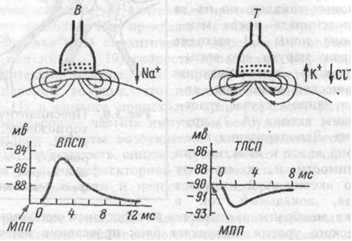 27) Аксошипиковые синапсы, их функциональная роль. Свойства химических синапсов (одностороннее проведение, синаптическая задержка и др.). Электрические синапсы, механизмы передачи возбуждения. Структурно-функциональное развитие нейронов в грудном и детском возрастах. Рост объема серого вещества мозга у детей. Свойства химических синапсов 1. Односторонняя проводимость — одно из важнейших свойств химического синапса. Асимметрия — морфологическая и функциональная — является предпосылкой для суще-ствования односторонней проводимости. 2. Наличие синаптической задержки: для того, чтобы в ответ на генерацию ПД в области пресинапса выделился медиатор и произошло изменение постсинаптического потенциала (ВПСП или ТПСП), требуется определенное время (синаптическая задержка). В среднем оно равно 0,2—0,5 мс. Это очень короткий промежуток времени, но когда речь идет о ре-флекторных дугах (нейронных сетях), состоящих из множества нейронов и синаптических связей, это латентное время суммируется и превращается в ощутимую величину — 300— 500 мс. В ситуациях, встречающихся на автомобильных дорогах, это время оборачивается трагедией для водителя или пешехода. 3. |