коллоквиум нормальная физиология. коллок нф 1. Физиология, ее предмет, роль задачи и формирование врачебной деятельности. Связь физиологии с другими науками
 Скачать 1 Mb. Скачать 1 Mb.
|

|
Возбудимость – это способность возбудимых тканей на действие раздражителя отвечать возбуждением, которое проявляется в виде биоэлектрического процесса и специфической ответной реакции. Подпороговое пассивное поведение клеточной мембраны называется электротоническим, или электротоном. Закон физиологического электротона(2 закон Пфлюгера) – закон об изменении физиологических свойств тканей при прохождении через них постоянного тока: При прохождении через ткань постоянного тока в области катода возбудимость и проводимость повышаются, а лабильность снижается(физиологический Кат-электротон), в области анода возбудимость и проводимость понижается, а лабильность повышается – физиологический АН-электротон. Дополнение Вериго: при прохождении через ткань сильного или длительного постоянного тока повышенная возбудимость в области катода сменяется пониженной – катодическая депрессия, а пониженная возбудимость в области анода сменяется повышенной – анодическая экзальтация. ИТОГ: Возбудимость клетки не зависит ни от величины мембранного потенциала, ни от величины критического уровня деполяризации, а определяется их соотношением: чем меньше между ними разница, тем выше возбудимость и наоборот. Закон Дюбуа-Раймона: Раздражающее действие постоянного тока не зависит ни от силы тока (его плотности), ни от длительности действия(времени), а определяется скоростью его нарастания: чем быстрее меняется ток во времени, тем большей раздражающей силой он обладает. Б) Пороговый потенциал (AV) — это минимальная величина, на которую надо уменьшить мембранный потенциал покоя, чтобы вызвать возбуждение (ПД). AV и возбудимость клеток находятся в обратных соотношениях: небольшая величина AV свидетельствует о высокой возбудимости клетки. Если, например, уменьшение мембранного потенциала (частичная деполяризация) на 5—10 мВ вызывает возникновение ПД, то возбудимость клетки высока. Напротив, большой AV (30—40 мВ) свидетельствует о более низкой возбудимости клетки. Однако во всех случаях ПД возникает только при достижении критического уровня деполяризации клеточной мембраны (Екр). В) Пороговая сила — это наименьшая сила раздражителя, способная вызвать возбуждение (ПД) при неограничении ее действия во времени (рис. 4.8). Сила раздражителя — понятие собирательное, оно отражает степень выраженности раздражающего воздействия стимула на ткань. Например, сила электрического тока выражается в амперах (А), температура среды — в градусах Цельсия (°С), концентрация химического вещества — в миллимолях на 1 л (ммоль/л), сила звука — в децибелах (дБ) и т.д. Г) Пороговое время — это минимальное время, в течение которого должен действовать на ткань раздражитель пороговой силы, чтобы вызвать ее возбуждение (см. рис. 4.8 — проекция точки А на абсциссу). Пороговое время называют также полезным временем, так как раздражитель обеспечивает деполяризацию только до критического уровня (Екр.). Далее ПД развивается независимо от действия раздражителя, дальнейшее раздражение уже становится ненужным — бесполезным. В эксперименте в клинической практике для оценки свойств возбудимой ткани чаще используют не пороговое время, а хронаксию. Это связано с тем, что определение порогового времени затрудненоРеобаза – это минимальная сила раздражителя, вызывающая возбуждение при его действии в течение неограниченно долгого времени. На практике порог и реобаза имеют одинаковый смысл. Чем ниже порог раздражения или меньше реобаза, тем выше возбудимость ткани. Хронаксия – это минимальное время действия раздражителя силой в две реобазы, необходимое для возникновения возбуждения. Этот параметр предложил рассчитывать Л.Лапик, для более точного определения показателя времени на кривой силы-длительности. Чем короче полезное время или хронаксия, тем выше возбудимость, и наоборот. В медицинской практике чаще всего определяется хронаксия мышц и двигательных нервов. Исследуется также хронаксия и чувствительной сферы. Хронаксия скелетных мышц человека колеблется от 0,1 до 0,7 мс. Хронаксия сгибателей у человека в 1,5-2 раза меньше хронаксии разгибателей. Хронаксиметрия — метод, определяющий величину хронаксии, т. е. наименьшего времени, в течение которого раздражитель удвоенной пороговой силы вызовет процесс возбуждения. Для измерения хронаксии пользуются специальным прибором — хронаксиметром, состоящим из источника постоянного тока, набора сопротивлений и приспособлений для дозировки времени действия тока, подающегося на объект. Хронаксиметрия применяется для определения дегенерации нерва при травмах различных нервных центров. Исследования хронаксии помогают установить сдвиги возбудимости при воздействии различных факторов: работы, тепла, холода, атмосферного давления и т.д. 18) Потенциал действия, понятие и схема. Фазы потенциала действия, их ионные механизмы. Функциональная роль потенциала действия. А) Потенциал действия – это электрофизиологический процесс, который выражается в быстром колебании мембранного потенциала вследствие изменения проницаемости мембраны и диффузии ионов в клетку и из клетки. 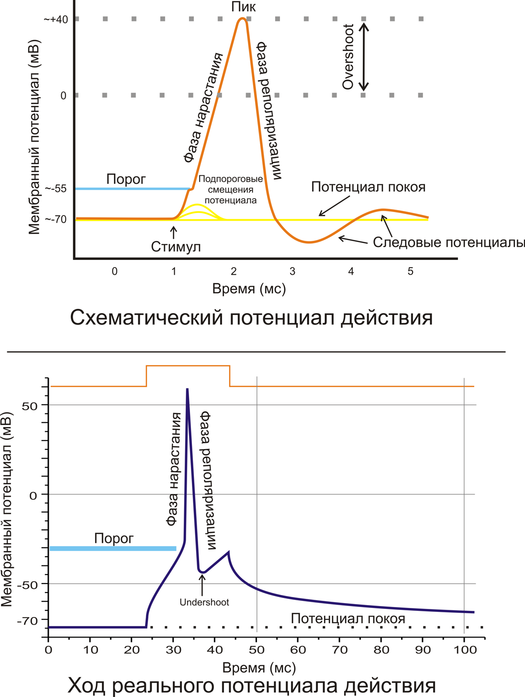 Б) Фаза деполяризации. При действии раздражителя на клетку начальная частичная деполяризация клеточной мембраны происходит без изменения ее проницаемости для ионов. Когда деполяризация достигает примерно 50% пороговой величины, возрастает проницаемость мембраны для Na+, причем в первый момент сравнительно медленно. В этот период движущей силой, обеспечивающей движение Na+ в клетку, являются концентрационный и электрический градиенты. Вспомним, что клетка внутри заряжена отрицательно (разноименные заряды притягиваются), а концентрация Na+ вне клетки в 12 раз больше, чем внутри клетки. Условием, обеспечивающим дальнейший вход Na+ в клетку, является увеличение проницаемости клеточной мембраны, который определяется состоянием воротного механизма натриевых каналов. Воротный механизм натриевых каналов расположен на внешней и внутренней стороне клеточной мембраны, воротный механизм калиевых каналов – только на внутренней стороне мембраны. В каналах для натрия имеются активационные m-ворота, которые расположены с внешней стороны клеточной мембраны, и инактивационные h-ворота, расположенные с внутренней стороны мембраны. В условиях покоя активационные m-ворота закрыты, инактивационные h-ворота открыты. Калиевые активационные ворота закрыты, а инактивационных калиевых ворот нет. Когда деполяризация клетки достигает критической величины, которая обычно составляет 50 мВ, проницаемость мембраны для Na+ резко возрастает, так как открывается большое количество потенциалзависимых m-ворот натриевых каналов и ионы натрия лавиной устремляются в клетку. Развивающаяся деполяризация клеточной мембраны вызывает дополнительное увеличение ее проницаемости и, соответственно, проводимости натрия: открываются все новые и новые активационные m-ворота. В итоге ПП исчезает, то есть становится равным нулю. Фаза деполяризации на этом заканчивается. Ее длительность составляет примерно 0,2-0,5 мс. Фаза инверсии. Процесс перезарядки мембраны представляет собой вторую фазу ПД – фазу инверсии. Фаза инверсии делится на восходящую и нисходящую составляющие. Восходящая часть. После исчезновения ПП вход в клетку ионов натрия продолжается, так как натриевые активационные m-ворота еще открыты. В результате заряд внутри клетки становится положительным, а снаружи-отрицательным. В течение доли миллисекунды ионы натрия еще продолжают входить в клетку. Таким образом, вся восходящая часть пика ПД обеспечивается в основном входом Na+ в клетку. Нисходящая составляющая фазы инверсии. Примерно через 0,2-0,5 мс после начала деполяризации рост ПД прекращается в результате закрытия натриевых инактивационных h-ворот и открытия калиевых активационных ворот. Поскольку калий находится преимущественно внутри клетки, он, согласно концентрационному градиенту, начинает быстро выходить из нее, вследствие чего уменьшается число положительно заряженных ионов в клетке. Заряд клетки снова начинает уменьшаться. Во время нисходящей составляющей фазы инверсии выходу ионов калия из клетки способствует также и электрический градиент. К+ выталкивается положительным зарядом из клетки и притягивается отрицательным зарядом снаружи клетки. Так продолжается до полного исчезновения положительного заряда внутри клетки. Калий выходит из клетки не только по управляемым каналам, но и по неуправляемым каналам – каналам утечки. Амплитуда ПД складывается из величины ПП и величины фазы инверсии, составляющей у разных клеток 10-50 мВ. Фаза реполяризации. Пока активационные калиевые каналы открыты, K+ еще продолжает выходить из клетки, согласно химическому градиенту. Заряд внутри клетки становится отрицательным, а снаружи – положительным, следовательно, электрический градиент резко тормозит выход ионов калия из клетки. Но так как сила химического градиента больше силы электрического градиента, ионы калия продолжают очень медленно выходить из клетки. Затем активационные калиевые ворота закрываются, остается только выход ионов калия по каналам утечки, то есть по концентрационному градиенту через неуправляемые каналы. Таким образом, ПД вызывается циклическим процессом поступления ионов натрия в клетку и последующего выхода калия из нее. Роль Са2+ в возникновении ПД в нервных клетках незначительна. Однако Са2+ играет очень важную роль в возникновении ПД сердечной мышцы, в передаче импульсов от одного нейрона к другому, от нервного волокна к мышечному, в обеспечении мышечного сокращения. Вслед за ПД возникают следовые явления (характерные для нейронов) – сначала следовая гиперполяризация, а затем следовая деполяризация. Следовая гиперполяризация клеточной мембраны обычно является следствием еще сохраняющейся повышенной проницаемости мембраны для ионов калия. Следовая деполяризация связана с кратковременным повышением проницаемости мембраны для Na+ и входом его в клетку согласно химическому и электрическому градиентам. Кроме этого существуют: а) так называемая фаза абсолютной рефрактерности, или полная невозбудимость клетки. Она приходится на пик ПД и продолжатся 1-2 мс; б) фаза относительной рефрактерности – период частичного восстановления клетки, когда сильное раздражение может вызвать новое возбуждение. Относительная рефрактерность соответствует конечной части фазы реполяризации и следовой гиперполяризации клеточной мембраны. В нейронах вслед за гиперполяризацией возможна частичная деполяризация клеточной мембраны. В этот период очередной потенциал действия можно вызвать более слабым раздражением, так как МП несколько меньше обычного. Этот период называется фазой экзальтации (период повышенной возбудимости). В) Роль ПД заключается в обеспечении передачи сигналов между нервными клетками, нервными центрами и рабочими органами, в мышцах ПД обеспечивает процесс электромеханического сопряжения. ПД подчиняется закону «всё или ничего». Если сила раздражения мала, то возникает локальный потенциал, который не распространяется. 19) Изменения возбудимости при возбуждении. Фазы возбудимости и их соотношение с фазами потенциала действия. Рефрактерность, ее механизмы. Изменение возбудимости при возбуждении. При возбуждении возбудимость изменяется пофазно. 1) фаза первичной экзальтации - возбудимость выше нормы, реакция на порошковый и подпороговый раздражитель (соответствует 1 фазе ПД - медленной деполяризации); 2) фаза абсолютной рефрактерности - ответная реакция на раздражитель отсутствует, что обусловлено инактивацией натриевых каналов (соответствует быстрой деполяризации ПД); 3) фаза относительной рефрактерности - возбудимость восстанавливается, и ответная реакция становится возможной только при действии раздражителя надпороговой силы, что обусловлено выходящим калиевым током (соответствует фазе реполяризации); 4) фаза вторичной экзальтации - ответная реакция на подпороговый раздражитель (соответствует следовой деполяризации); 5) фаза субнормальной возбудимости - возбудимость ниже нормы, ответ возможен на действие надпороговой силы (соответствует следовой гиперполяризации) 20) Законы раздражения возбудимых тканей: закон силы (силовых отношений), пессимум силы раздражителя, закон «все или ничего», закон крутизны нарастания силы раздражителя (аккомодация). Закон "все или ничего": При допороговых раздражениях клетки в ткани ответной реакции не возникает. При пороговой силе раздражителя развивается максимальная ответная реакция, поэтому увеличение силы раздражения выше пороговой не сопровождается ее усилением. В соответствии с этим законом реагирует на раздражения одиночное нервное и мышечное волокно, сердечная мышца.Закон силы: Чем больше сила раздражителя, тем сильнее ответная реакция. Однако выраженность ответной реакции растет лишь до определенного максимума. Закону силы подчиняется целостная скелетная, гладкая мышца, так как они состоят из многочисленных мышечных клеток, имеющих различную возбудимость. Закон длительности: Между силой и длительностью действия раздражителя имеется определенная взаимосвязь. Чем сильнее раздражитель, тем меньшее время требуется для возникновения ответной реакции. Зависимость между пороговой силой и необходимой длительностью раздражения отражается кривой силы длительности. По этой кривой можно определить ряд параметров возбудимости. Закон градиента или аккомодации: реакция ткани на раздражение зависит от его градиента, т.е. чем быстрее нарастает сила раздражителя во времени тем быстрее возникает ответная реакция. При низкой скорости нарастания силы раздражителя растет порог раздражения. Поэтому если сила раздражителя, возрастает очень медленно возбуждения не будет. Это явление называется аккомодацией. Для возникновения возбуждения сила раздражающего тока должна нарастать достаточно круто. Величина ответной реакции возбудимых тканей прямопропорциональна (в определенных пределах) силе раздражителя эта зависимость выражается уравнением: В = 1/S, где В - возбудимость, S - сила раздражителя. Однако не только сила раздражителя, но и время (длительность) его воздействия оказывает влияние на величину ответной реакции. Более того, пороговый раздражитель при более коротком времени воздействия может не вызвать ответной реакции, т.е. оказаться допороговым. Представленная на (рис. 4) кривая называется кривой силы - длительности, или силы - времени. Она была получена при исследовании возбудимости нервов и мышц Гоорвегом (1892), Вейсом (1901) и Лапиком (1909). |