СРС-3(реферат) Нуралибеков и Давронова. Генетическое сцепление и картирование генов человека
 Скачать 92.34 Kb. Скачать 92.34 Kb.
|

|
МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ РЕСПУБЛИКИ КАЗАХСТАН КАЗАХСКИЙ НАЦИОНАЛЬНЫЙ УНИВЕРСИТЕТ ИМЕНИ АЛЬ-ФАРАБИ  Факультет: биологии и биотехнологии Кафедра: биотехнологии «РЕФЕРАТ» На тему: ГЕНЕТИЧЕСКОЕ СЦЕПЛЕНИЕ И КАРТИРОВАНИЕ ГЕНОВ ЧЕЛОВЕКА. Выполнили : студенты 3-курса (мед.бт.) Нуралибеков С.Ш. Давронова М.А. Проверила : к.б.н. ,доцент кафедры молекулярной биологии и генетики Омирбекова Н.Ж. АЛМАТЫ 2018 СОДЕРЖАНИЕ Картирование и определение первичной структуры генома человека…………….….3 Генетические карты сцепления…………………………………………………………..3 Современные методы построения генетических карт сцепления……..........……...….5 ПЦР в исследованиях генома человека………………………………....………….……8 Физические карты низкого разрешения…………………………………………..….….9 Физические карты высокого разрешения……………..………………………..………11 Список использованных источников ………………...……………..………………….13 Картирование и определение первичной структуры генома человека После краткого рассмотрения основных методов, наиболее часто используемых в молекулярной генетике для исследования структуры и механизмов функционирования генов, представляется целесообразным на примере генома человека подробнее познакомиться с практическим применением этих методов и их модификаций для изучения больших геномов. В целях всестороннего исследования генома человека, этого колоссального по объему хранилища его генетической информации, недавно была разработана и воплощается в жизнь специальная международная программа "Геном человека" ("Human Genome Project"). Основной задачей программы является построение исчерпывающих генетических карт большого разрешения каждой из 24 хромосом человека, которое, в конечном счете, должно завершиться определением полной первичной структуры ДНК этих хромосом. В настоящее время работы по проекту идут полным ходом. В случае успешного его завершения (а это по планам должно произойти в 2003 г.) у человечества появятся перспективы досконального изучения функциональной значимости и механизмов функционирования каждого из его генов, а также генетических механизмов, управляющих биологией человека, и установления причин большинства патологических состояний его организма. Основные подходы к картированию генома человека Решение основной задачи программы "Геном человека" включает три основных этапа. На первом этапе необходимо специфическим образом разделить каждую индивидуальную хромосому на части меньшего размера, позволяющего их дальнейший анализ известными методами. Вторая стадия исследований предполагает определение взаимного расположения этих индивидуальных фрагментов ДНК друг относительно друга и их локализации в самих хромосомах. На завершающем этапе необходимо произвести собственно определение первичной структуры ДНК каждого из охарактеризованных фрагментов хромосом и составить полную непрерывную последовательность их нуклеотидов. Решение задачи не будет полным, если в найденных последовательностях нуклеотидов не удастся локализовать все гены организма и определить их функциональное значение. Прохождение трех вышеперечисленных этапов требуется не только для получения исчерпывающих характеристик генома человека, но и любого другого генома большого размера. Генетические карты сцепления Генетические карты сцепления представляют собой одномерные схемы взаимного расположения генетических маркеров на индивидуальных хромосомах. Под генетическими маркерами понимают любые наследуемые фенотипические признаки, различающиеся у отдельных особей. Фенотипические признаки, отвечающие требованиям генетических маркеров, весьма разнообразны. Они включают в себя как особенности поведения или предрасположенность к определенным заболеваниям, так и морфологические признаки целых организмов или их макромолекул, различающихся по структуре. С развитием простых и эффективных методов исследования биологических макромолекул такие признаки, известные под названием молекулярных маркеров, стали наиболее часто использоваться при построении генетических карт сцепления. Прежде чем перейти к рассмотрению методов построения таких карт и их значения для исследования генома, необходимо напомнить, что термин "сцепление" употребляется в генетике для обозначения вероятности совместной передачи двух признаков от одного из родителей потомству. При образовании половых клеток (гамет) у животных и растений на стадии мейоза, как правило, происходит синапсис (конъюгация) гомологичных хромосом. Сестринские хроматиды гомологичных хромосом соединяются по всей длине друг с другом, и в результате кроссинговера (генетической рекомбинации между хроматидами) происходит обмен их частями. Чем дальше два генетических маркера располагаются друг от друга на хроматиде, тем больше вероятность того, что разрыв хроматиды, необходимый для кроссинговера, произойдет между ними, и два маркера в новой хромосоме, принадлежащей новой гамете, окажутся отделенными друг от друга, т.е. их сцепление нарушится. Единицей сцепления генетических маркеров является морганида (единица Моргана, М), которая содержит 100 сантиморганид (сМ). 1 сМ соответствует физическому расстоянию на генетической карте между двумя маркерами, рекомбинация между которыми происходит с частотой 1%. Выраженная в парах оснований 1 сМ соответствует 1 млн п.о. (м.п.о.) ДНК. Генетические карты сцепления правильно отражают порядок расположения генетических маркеров на хромосомах, однако полученные при этом значения расстояний между ними не соответствуют реальным физическим расстояниям. Обычно данный факт связывают с тем, что эффективность рекомбинации между хроматидами на отдельных участках хромосом может сильно различаться. В частности, она подавлена в гетерохроматиновых участках хромосом. С другой стороны, в хромосомах часто встречаются "горячие точки" рекомбинации. Использование частот рекомбинации для построения физических генетических карт без учета этих факторов будет приводить к искажениям (соответственно занижению или завышению) реальных расстояний между генетическими маркерами. Таким образом, генетические карты сцепления являются наименее точными из всех имеющихся типов генетических карт, и их можно рассматривать только в качестве первого приближения к реальным физическим картам. Тем не менее, на практике именно они и только они позволяют локализовать сложные генетические маркеры (например ассоциированные с симптомами заболевания) на первых этапах исследования и дают возможность их дальнейшего изучения. Необходимо помнить, что в отсутствие кроссинговера все гены, находящиеся на индивидуальной хромосоме, передавались бы от родителей потомству вместе, поскольку они физически сцеплены друг с другом. Поэтому индивидуальные хромосомы образуют группы сцепления генов, и одной из первых задач построения генетических карт сцепления является отнесение исследуемого гена или последовательности нуклеотидов к конкретной группе сцепления. В след. таблице перечислены современные методы, которые, по данным В.А. МакКьюзика, наиболее часто использовались для построения генетических карт сцепления до конца 1990 г. Современные методы построения генетических карт сцепления
Гибридизация соматических клеток. Одним из наиболее популярных методов отнесения генетического маркера (функционально активного гена) к конкретной группе сцепления является гибридизация (слияние друг с другом) соматических клеток разных биологических видов организмов, один из которых – исследуемый. У межвидовых гибридов соматических клеток в процессе культивирования происходит утрата хромосом преимущественно одного из биологических видов. Потеря хромосом носит, как правило, случайный характер, и образующиеся клоны клеток содержат оставшиеся хромосомы в разных сочетаниях. Анализ клонов, содержащих разные наборы хромосом исследуемого вида, позволяет определить, с какой из этих оставшихся хромосом ассоциирована экспрессия исследуемого маркера, и, следовательно, локализовать ген на конкретной хромосоме. Гибридизация in situ. Метод гибридизации in situ также широко используется для картирования последовательностей нуклеотидов на хромосомах. С этой целью препараты фиксированных хромосом гибридизуют (инкубируют при повышенной температуре с последующим охлаждением) с исследуемыми последовательностями нуклеотидов, меченными радиоактивной, флуоресцентной или иной меткой. После отмывания несвязавшейся метки оставшиеся меченые молекулы нуклеиновых кислот оказываются ассоциированными с участками хромосом, содержащими последовательности, комплементарные исследуемым меченым последовательностям нуклеотидов. Полученные гибриды анализируют с помощью микроскопа либо непосредственно, либо после авторадиографии. Для этой группы методов характерна более высокая разрешающая способность, чем для гибридизации соматических клеток, поскольку они позволяют локализовать изучаемые последовательности нуклеотидов на хромосомах. По мере выполнения программы "Геном человека" в руках исследователей появляется все больше изолированных последовательностей нуклеотидов, которые можно использовать в качестве зондов для гибридизации in situ. В связи с этим данные методы по частоте использования в последнее время прочно выходят на первое место. Наиболее популярной оказывается группа методов, получивших название флуоресцентной гибридизации in situ (fluorescence in situ hybridization – FISH), при проведении которой используются полинуклеотидные зонды, содержащие флуоресцентную метку. В частности, в 1996 г. было опубликовано >600 работ, в которых описано использование этого метода. Семейный генетический анализ сцепления. Эта группа методов часто используется в медицинской генетике для выявления связи (сцепления) между симптомами заболевания, вызываемого мутацией в неизвестном гене, и другими генетическими маркерами. В данном случае в качестве одного из генетических маркеров выступают сами симптомы заболевания. В геноме человека обнаружено большое количество полиморфизмов, в том числе ПДРФ. ПДРФ распределены более или менее равномерно в геноме человека на расстоянии 5–10 сМ друг от друга. Чем ближе индивидуальные полиморфные локусы расположены к гену, ответственному за заболевание, тем меньше вероятность их разделения при рекомбинации в мейозе и тем чаще они будут встречаться вместе у больного индивидуума и вместе передаваться от родителей потомству. Клонировав протяженный участок генома, включающий соответствующий полиморфный маркер (его отбор из клонотеки геномной ДНК проводят с помощью зонда), можно одновременно вместе с ним с большой вероятностью выделить ген, вызывающий наследственное заболевание. Такие подходы были, в частности, успешно применены для проведения семейного анализа и выделения соответствующих генов при мышечной дистрофии Дюшенна, кистозном фиброзе почек (муковисцидозе) и миотонической дистрофии. Информативность отдельных ПДРФ генома человека зависит от уровня их гетерозиготности в исследуемой популяции. Мерой информативности ПДРФ как генетического маркера по предложению Д. Ботштейна и соавторов (1980 г.) принято считать значение содержания полиморфной информации PIC (polymorphism information content), которое представляет собой отношение числа скрещиваний, в которых хотя бы у одного из родителей исследуемый полиморфный маркер находится в гетерозиготном состоянии, ко всем скрещиваниям. Определение эффекта дозы гена и использование хромосомных аберраций. Этими методами обнаруживают корреляции между уровнем экспрессии исследуемого гена и количеством конкретных хромосом в анеуплоидных линиях клеток или структурными перестройками хромосом (хромосомными мутациями – аберрациями). Анеуплоидией называют наличие у клетки, ткани или целого организма числа хромосом, не равного типичному для данного биологического вида. Хромосомные аберрации в виде транслокаций участков хромосом в гетерохроматиновые области тех же самых или других хромосом часто сопровождаются подавлением транскрипции генов, расположенных в транслоцированных участках или в хромосоме-акцепторе (мозаичный эффект положения). Использование синтении. Синтения – это структурное сходство групп сцепления генов у организмов разных биологических видов. В частности, в геномах человека и мыши известно несколько десятков синтеничных групп генов. Наличие феномена синтении позволяет суживать круг поиска места локализации исследуемого гена на хромосомах, ограничивая его областью известных генов, принадлежащих к конкретной синтеничной группе. Сегрегация генов, индуцируемая ионизирующим излучением. С помощью этого метода определяют расстояние между исследуемыми генами путем оценки вероятности их разделения (сегрегации) после облучения клеток определенной стандартной дозой ионизирующего излучения. Облученные клетки спасают от гибели гибридизацией с соматическими клетками грызунов, и у соматических гибридов в культуре определяют наличие исследуемых маркеров облученных клеток. В итоге удается сделать вывод о наличии или отсутствии сцепления (физическом расстоянии) между этими генами. Среди других методов следует упомянуть способы, основанные на использовании для картирования генов больших фрагментов ДНК, образуемых под действием крупнощепящих рестриктаз. После расщепления геномной ДНК образующиеся фрагменты разделяют электрофорезом в импульсном электрическом поле и далее их гибридизуют по Саузерну с зондами, соответствующими картируемым генам. Если после проведения гибридизации сигналы обоих зондов локализуются на одном и том же крупном фрагменте ДНК, это говорит о тесном сцеплении таких генов. ПЦР в исследованиях генома человека Полимеразная цепная реакция занимает центральное место в разработке подходов к практическому осуществлению программы "Геном человека". Как уже обсуждалось выше, с помощью ПЦР можно быстро и эффективно амплифицировать почти любой короткий участок генома человека, и полученные продукты ПЦР далее использовать в качестве зондов для картирования соответствующих участков на хромосомах путем гибридизации по Саузерну или in situ. Концепция STS. Одной из ключевых концепций, лежащих в основе картирования генов человека в рамках обсуждаемой программы, является концепция сайтов, привязанных к последовательностям (sequence-tagged sites – STS). В соответствии с этой концепцией все фрагменты ДНК, используемые для построения генетических или физических карт, можно однозначно идентифицировать с помощью последовательности нуклеотидов длиной в 200–500 п.о., которая будет уникальной для данного фрагмента. Каждый из этих сайтов необходимо секвенировать, что даст возможность в дальнейшем их амплифицировать с помощью ПЦР и применять в качестве зондов. Использование STS позволило бы применять их последовательности в виде продуктов ПЦР в качестве зондов для направленного выделения любого фрагмента ДНК того или иного участка генома из клонотек геномных последовательностей. В результате могут быть созданы базы данных, включающие локализацию и структуру всех STS, а также праймеров, необходимых для их амплификации. Это избавило бы лаборатории от необходимости хранения многочисленных клонов и их рассылки в другие лаборатории для проведения исследований. Кроме того, STS создают основу для разработки единого языка, на котором разные лаборатории могли бы описывать свои клоны. Таким образом, конечным результатом разработки концепции STS была бы исчерпывающая карта STS генома человека. Теоретически для построения генетической карты размером в 1 сМ необходимо 3000 полностью информативных, полиморфных ДНК-маркеров. Однако поскольку полиморфные маркеры распределены в геноме неравномерно и лишь немногие из них полностью информативны, реальное число маркеров, требуемых для построения карты такого размера, оценивается в 30–50 тысяч. Для получения маркеров, соответствующих исследуемым участкам хромосом, в настоящее время часто применяют праймеры, соответствующие диспергированным повторяющимся последовательностям, среди которых первыми стали использовать Alu-последовательности . Alu-ПЦР. Диспергированные повторяющиеся Alu-последовательности характерны именно для генома человека. Праймеры, специфичные в отношении Alu-последовательностей, используют для амплификации участков ДНК генома человека, заключенных между Alu-повторами, которые располагаются в среднем на расстоянии 4–10 т.п.о. друг от друга. Другим вариантом Alu-ПЦР является направленный синтез с ее помощью ДНК-зондов к участкам хромосом, полученным после лазерной фрагментации, индивидуальным хромосомам, выделенным с помощью проточной цитофлуориметрии, или ДНК гибридных клеток, содержащих определенную часть генома человека. Кроме того, Alu-ПЦР используют для получения уникальных фингерпринтов, характеризующих клеточные гибриды с точки зрения стабильности их генома, а также для характеристики фрагментов ДНК человека, клонированных в YAC-векторах, космидах или векторах на основе ДНК бактериофагов. Уникальность Alu-последовательностей для генома человека делает возможным их применение для "прогулок по хромосомам" , а также для расширения существующих контигов. Поскольку в геноме человека >90% умеренно повторяющихся последовательностей представлены семействами Alu и KpnI, неудивительно, что последние также применяются в ПЦР для тех же целей, что и Alu. Однако здесь профили продуктов ПЦР менее сложны, поскольку последовательности KpnI повторяются в геноме реже и обладают характерной локализацией в хромосомах. ПЦР активно используется для выявления полиморфных молекулярных маркеров при построении генетических карт сцепления, основные принципы получения которых были рассмотрены выше. Этот метод оказывается полезным и при секвенировании ДНК, а также при построении физических карт высокого разрешения для генома человека. О последних двух сферах применения ПЦР подробнее речь пойдет ниже. Физические карты низкого разрешения В отличие от рассмотренных выше генетических карт сцепления физические карты генома отражают реальное расстояние между маркерами, выражаемое в парах оснований. Физические карты различаются по степени их разрешения, т.е. по тем деталям структуры генома, которые на них представлены. Исчерпывающая физическая карта генома человека максимального разрешения будет содержать полную нуклеотидную последовательность всех его хромосом. На другом полюсе физических карт с минимальным разрешением находятся хромосомные (цитогенетические) карты генома. Четыре типа генетических карт геномной ДНК и их взаимоотношения 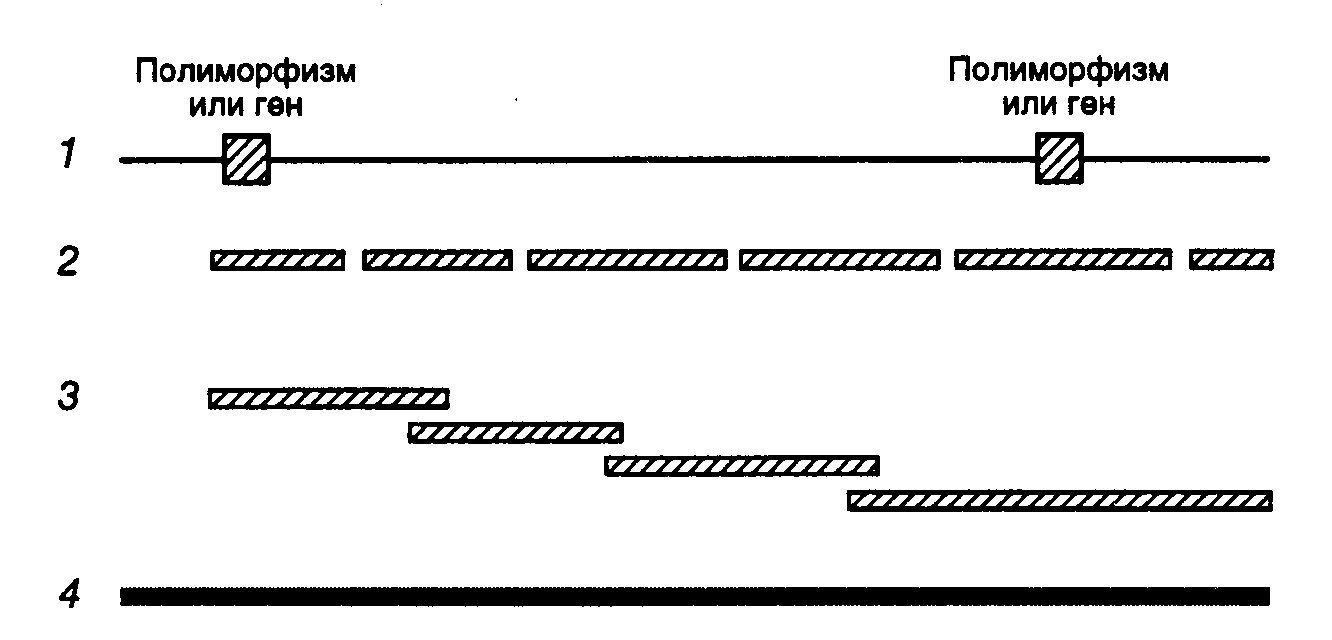 1 – генетическая карта сцепления, 2 – физическая рестрикционная карта, пробелы обозначают места расщепления ДНК рестриктазами, 3 – физическая карта контигов, показаны перекрывающиеся клоны ДНК, полученные с помощью YAC-векторов, 4 – исчерпывающая физическая карта в виде последовательности нуклеотидов ДНК. На всех картах представлен один и тот же участок хромосомы Хромосомные карты. Хромосомные карты генома человека получают локализацией генетических маркеров на индивидуальных хромосомах с использованием цитогенетических методов, включая авторадиографию и FISH. В последних двух случаях радиоактивная или флуоресцентная метки, ассоциированные с исследуемыми генетическими локусами интактных хромосом, выявляются с помощью световой микроскопии. Еще совсем недавно хромосомные карты позволяли локализовать исследуемый фрагмент ДНК на участке хромосомы протяженностью 10 м.п.о. Современные методы гибридизации in situ с использованием метафазных хромосом, главным образом, метод FISH, локализуют полинуклеотидные маркеры в пределах 2–5 м.п.о. Более того, при гибридизации in situ с интерфазными хромосомами, в которых генетический материал находится в менее компактной форме, разрешающая способность хромосомных карт приближается к 100 т.п.о. Точность хромосомных карт повышается и с использованием современных генетических методов. Например, способность ПЦР амплифицировать сегменты ДНК единичного сперматозоида позволяет исследовать большое число мейозов, как бы законсервированных в отдельных образцах спермы. В результате появляется возможность проверки взаимного расположения генетических маркеров, локализованных на хромосомных картах более грубыми методами. Карты кДНК. Карты кДНК отражают положение экспрессирующихся участков ДНК (экзонов) относительно известных цитогенетических маркеров (бэндов) на метафазных хромосомах. Поскольку такие карты дают представление о локализации транскрибирующихся участков генома, в том числе и генов с неизвестными функциями, они могут быть использованы для поиска новых генов. Этот подход особенно полезен при поиске генов, повреждения которых вызывают заболевания человека, в том случае если приблизительная локализация таких участков хромосом уже предварительно проведена на генетических картах сцепления в результате семейного генетического анализа. Физические карты высокого разрешения 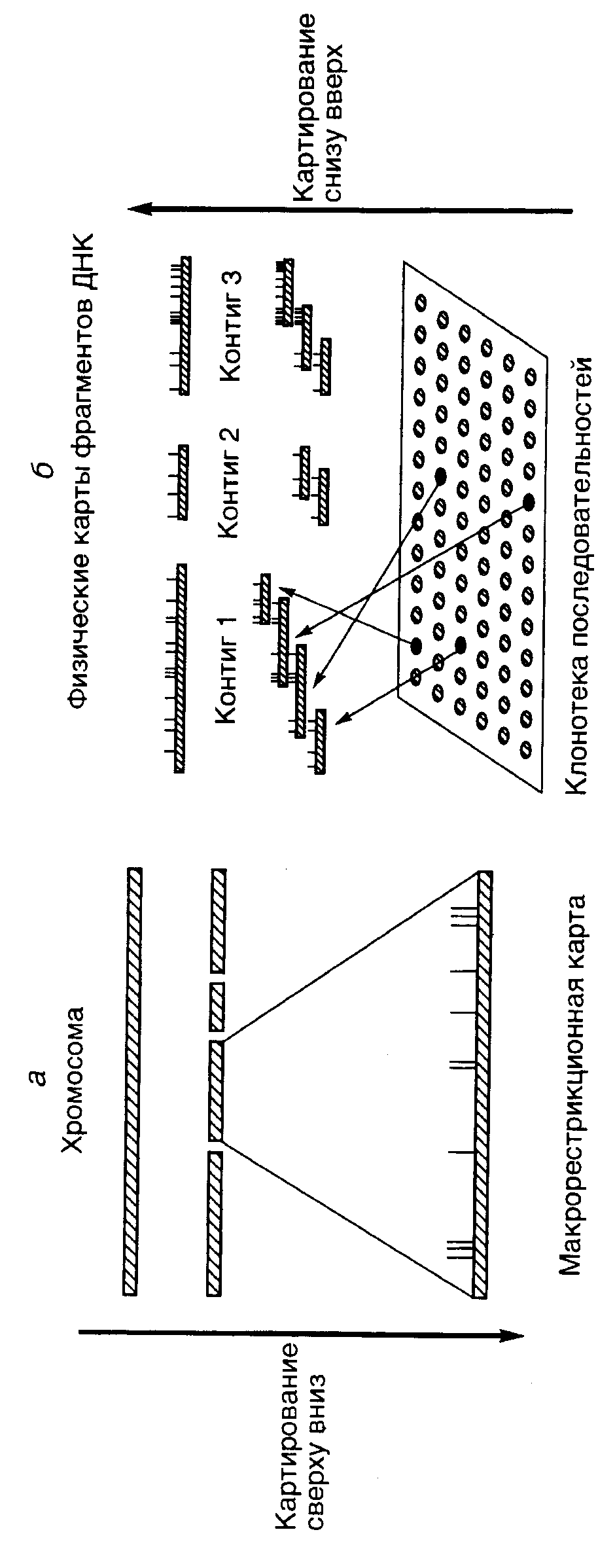 Две стратегии построения физических карт ДНК а – стратегия "сверху вниз": ДНК целой хромосомы расщепляется крупнощепящими рестриктазами, для каждого из индивидуальных фрагментов ДНК строится рестрикционная карта; б – стратегия "снизу вверх", индивидуальные YAC-клоны после идентификации объединяются в контиги В попытках построения карт генома человека высокого разрешения экспериментально реализуются два альтернативных подхода, получивших названия картирования сверху вниз (top-down mapping) и картирования снизу вверх (bottom-up mapping). При картировании сверху вниз исходным в анализе является препарат ДНК индивидуальной хромосомы человека. ДНК разрезается крупнощепящими рестриктазами (например NotI) на длинные фрагменты, которые после разделения электрофорезом в импульсном электрическом поле подвергаются дальнейшему рестрикционному анализу с другими рестриктазами. В результате получают макрорестрикционную карту, на которой достаточно полно представлены все последовательности исследуемой хромосомы или ее части, однако ее разрешение невысоко. На такой карте очень трудно локализовать индивидуальные гены. К тому же каждая индивидуальная карта редко охватывает протяженные сегменты ДНК (как правило, не более 1–10 м.п.о.). При картировании генома человека снизу вверх на основе препарата суммарной ДНК генома или индивидуальной хромосомы получают серию случайных клонов протяженных последовательностей ДНК (10–1000 т.п.о), часть из которых перекрывается друг с другом. В качестве вектора для клонирования в этом случае часто используют искусственные минихромосомы бактерий (BAC) или дрожжей (YAC), подробно описанные в разделе 7.2.4. Серия частично перекрывающихся и дополняющих друг друга клонов образует непрерывную состыкованную (contiguous) последовательность нуклеотидов ДНК, получившую название контига (contig). Правильность полученных контигов подтверждают гибридизацией in situ (FISH) с одновременной их привязкой к определенным участкам исследуемых хромосом. Карты, основанные на контигах, представляют полную информацию о структуре отдельных сегментов хромосом и позволяют локализовать отдельные гены. Однако такие карты трудно применять для реконструкции целых хромосом или протяженных их участков из-за отсутствия соответствующих клонов в имеющихся клонотеках генов. Основная проблема, которую приходится решать при использовании обоих подходов к построению физических карт высокого разрешения, – объединение разрозненных фрагментов ДНК в непрерывные последовательности нуклеотидов. Чаще всего для этого применяют специальные клонированные фрагменты ДНК, получившие название связующих (linking) клонов. Фрагменты ДНК из связующих клонов содержат в своих внутренних частях последовательности нуклеотидов крупнощепящих рестриктаз и, следовательно, представляют собой места стыковки фрагментов ДНК, используемых на первых этапах физического картирования. Гибридизацией по Саузерну, при проведении которой в качестве зондов используют фрагменты ДНК связующих клонов, определяют фрагменты ДНК физических карт, содержащие последовательности нуклеотидов окрестностей сайтов рестрикции крупнощепящих рестриктаз. Если два таких фрагмента найдены, то соответствующий связующий клон перекрывает оба этих фрагмента и является их частью. Связующие клоны, в свою очередь, отбирают из клонотек генов с помощью зондов, которые представляют собой последовательности нуклеотидов сайтов рестрикции крупнощепящих рестриктаз. СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ 1) Clark M.S. Comparative genomics: The key to understanding the Human Genome Project // BioEssays. 1999. Vol. 21. P. 21–30. 2) Billings P.R., Smith C.L., Cantor C.L. New techniques for physical mapping of the human genome // FASEB J. 1991. Vol. 5. P. 28–34. 3) Георгиев Г.П. Гены высших организмов и их экспрессия. М.: Наука, 1989. 254 с. 4) http://referatwork.ru/refs/source/ref-8543.html 5) https://studfiles.net/preview/5857530/page:173/ |