гормоны. Гормоны это биологически высокоактивные вещества, синтезирующиеся и выделяющиеся во внутреннюю среду организма эндокринными железами и оказывающие регулирующее влияние на функции удаленных от места их секреции органов и систем организма
 Скачать 157.5 Kb. Скачать 157.5 Kb.
|

Общая характеристика иерархических систем нейрогуморальной регуляцииВ поддержании физиологического уровня гормонов в крови участвует целый ряд механизмов гомеостаза, обеспечивающих точный обмен сигналами между гормон-секретирующей железой и тканью-мишенью, причем нередко это осуществляется при посредничестве одной или нескольких других эндокринных желез. Наиболее часто встречается механизм регуляции, основанный на отрицательной обратной связи. В особенности это свойственно системе гипоталамус — гипофиз — железа-мишень. Секреция всех гормонов передней доли гипофиза (соматотропина, тиротропина, кортикотропина, фоллитропина, лютропина, пролактина) находится под контролем веществ пептидной природы, образующихся в гипоталамусе и способных усиливать или уменьшать образование тропных гормонов гипофиза. Гипоталамус — область головного мозга, примыкающая к гипофизу. Именно в гипоталамусе происходит «переключение» нервных импульсов на гуморальные. К настоящему времени из гипоталамуса выделено несколько рилизинг-гормонов, или, как их еще иначе называют, либеринов: тиреотропин-рилизинг-гормон (тиролиберин, сокращенно ТРГ), гонадотропин-рилизинг-гормон (люлиберин), гормон, стимулирующий выработку кортикотропного гормона, — кортикотропин-рилизинг-гормон (кортиколиберин), гормон, вызывающий секрецию пролактина (пролактолиберин), меланоцит-освобождающий гормон и др. Среди гормонов гипоталамуса, оказывающих тормозящее действие на гипофизарную функцию, выделены (и синтезированы) вещества, названные статинами: соматостатин, пролактостатин, меланоцитостатин и др. По портальной системе кровообращения, связывающей сетью своих широко разветвленных капилляров срединное возвышение гипоталамуса с паренхимой аденогипофиза, либерины «спускаются» в переднюю долю гипофиза, вызывая усиленное образование и отделение в кровь тропных гормонов гипофиза. Под влиянием тропных гормонов гипофиза возрастает секреторная активность периферических эндокринных органов. Изменение концентрации метаболитов в клетках – мишенях по механизму отрицательной обратной связи может подавлять подавляет синтез гормонов, действуя на гипоталамус; синтез и секреция тропных гормонов подавляются гормонами периферических желез. В других случаях отрицательная обратная связь осуществляется с помощью отдельных метаболитов или субстратов, концентрация которых в плазме крови меняется при воздействии гормона на ткань-мишень. Например, увеличение концентрации глюкозы в крови (гипергликемия) вызывает измеряемое высвобождение инсулина, который усиливает потребление и утилизацию глюкозы в ряде тканей; в результате уровень глюкозы в крови возвращается к норме, что в свою очередь снижает секрецию инсулина. 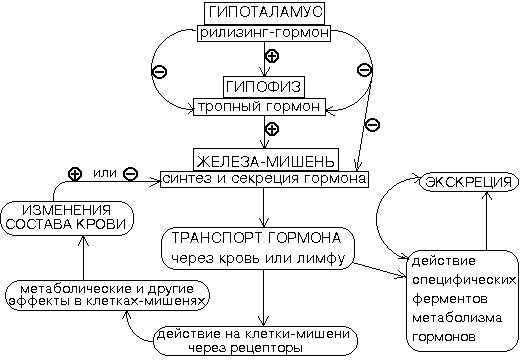 . .Рис. 1. Данная схема включает в себя высшие уровни регуляции в эндокринной системе - гипоталамус и гипофиз, вырабатывающие гормоны, которые сами влияют на процессы синтеза и секреции гормонов других эндокринных клеток. Из этой же схемы видно, что скорость синтеза и секреции гормонов может изменяться также под действием гормонов из других желез или в результате стимуляции негормональными метаболитами. Мы видим также наличие отрицательных обратных связей (-) - торможение синтеза и(или) секреции после устранения первичного фактора, вызвавшего ускорение продукции гормона. В результате содержание гормона в крови поддерживается на определенном уровне, который зависит от функционального состояния организма. Биосинтез гормонов Белково-пептидные гормоны чаще всего синтезируются в виде неактивных предшественников и образуются по схеме препрогормон прогормонактивный гормон. Препрогормон синтезируется на рибосоме. Препрогормон имеет на N- конце сигнальный пептид, необходимый для переноса молекулы в шероховатый эндоплазматический ретикулум, а затем в комплекс Гольджи. После удаления сигнального пептида препрогормон превращается в прогормон. В комплексе Гольджи прогормон пакуется в секреторные везикулы и непосредственно в них происходит дальнейший частичный протеолиз с образованием активного гормона. При синтезе гормонов из числа сложных белков гликопротеинов (например, фолликулостимулирующего (ФСГ) или тиреотропного (ТТГ) гормонов гипофиза) в процессе созревания происходит включение углеводного компонента в структуру гормона. Стероидные гормоны синтезируются из холестерина или из его производного - дегидрохолестерина. Общим предшественником всех кортикостероидов служит холестерин. При синтезе кортикостероидов образуется более 40 метаболитов, дающихся по структуре и биологической активности. Основными кортикостероидами, обладающими выраженной гормональной активностью, являются кортизол—глюкокортикоид, альдостерон—минералокортикоид и андрогены. На первом этапе синтеза кортикостероидов сходят превращение холестерина в прегненолон пугем отщепления 6-углеродного фрагмента от боковой цепи холестерина и окисление С20 углеродного атома. Прегненолон превращается в прогестерон -предшественник С21-стероидов — кортизола и альдостерона, и С19-стероидов, предшественники андрогенов. Каким именно стероидом окажется конечный продукт, зависит от набора ферментов в клетке и последовательности реакций гидроксилирования. Первичное гидроксилирование прогестерона 17 - гидроксилазой, а затем 21- и 11-гидроксилазой к синтезу кортизола. Реакции образования альдостерона включают гидроксилирование прогестерона сначала 21-гидроксилазой, а затем 11-гидроксилазой. Стероидные гормоны транспортируются кровью в комплексе со специфическими транспортными белками. Скорость синтеза и секреции кортизола регулируется гипоталамо-гипофизарной системой по механизму отрицательной обратной связи. Катаболизм гормонов коры надпочечников происходит прежде всего в печени. Здесь протекают реакции гидроксилирования, окисления и восстановления гормонов. Продукты катаболизма кортикостероидов (кроме кортикостерона и альдостерона) выводятся с мочой в форме 17-кетостероидов. Эти продукты метаболизма выделяются преимущественно в виде конъюгатов с глюкуроновой и серной кислотами. У мужчин 2/3 кетостероидов образуется за счет кортикостероидов и 1/3 — за счет тестостерона (всего 12—17 мг/сут). У женщин 17-кетостероиды образуются преимущественно за счет кортикостероидов (7—12 мг/сут). Биосинтез катехоламинов и дофамина. Исходным соединением является тирозин. Ключевым ферментом синтеза катехоламинов является тирозингидроксилаза, катализирующая превращение тирозина в ДОФА (L--(3,4 дигидроксифенил)- -аланин. Его превращение в дофамин катализирует декарбоксилаза ароматической аминокислоты. Превращение дофамина в норадреналин катализирует фермент дофамин - -гидроксилаза. Превращение норадреналина в адреналин катализирует фермент норадреналин –N- метилтрансфераза (фенилаэтаноламин N- метилтрансфераза). После окончания синтеза катехоламины и дофамин в свободном состоянии пакуются в специальные секреторные гранулы. Биосинтез йодтиронинов стимулируется ТТГ гипофиза. Йодтиронины синтезируются в составе белка тироглобулина. Тироглобулин — гликопротеин, синтезируется в базальной части клетки и хранится во внеклеточном коллоиде, где происходит йодирование остатков тирозина и образование йодтиронинов. Под действием тиропероксидазы окисленный иод реагирует с остатками тирозина с образованием монойодтиронинов (МИТ) и дийодтиронинов (ДИТ) Две молекулы ДИТ конденсируются с образованием тироксина (Т4), а МИТ и ДИТ с образованием йодтиронина (Тз). Йодтиреоглобулин транспортируется в клетку путем эндоцитоза и гидролизуется ферментами лизосом с освобождением Т3 и Т4. В крови йодтиронины находятся в связанной форме в комплексе с тироксинсвязывающим белком. Только 0,03% Т4 и 0,3% Т3 находятся в свободном состоянии. Биологическая активность йодтиронинов обусловлена несвязанной фракцией. Транспортные белки служат своеобразным депо, которое может обеспечить дополнительное количество свободных гормонов. Синтез и секреция йодтиронинов регулируются гипоталамо-гипофизарной системой. Гормоны передней доли гипофиза В аденогипофизе образуются следующие гормоны: адренокортикотропный (АКТГ), или кортикотропин; тиреотропный (ТТГ), или тиреотропин, гонадотропные: фолликулостимулирующий (ФСГ), или фоллитропин, и лютеинизирующий (ЛГ), или лютропин, соматотропный (СТГ), или гормон роста, или соматотропин, пролактин. Первые 4 гормона регулируют функции так называемых периферических желез внутренней секреции. Соматотропин и пролактин сами действуют на ткани-мишени. Адренокортикотропный гормон (АКТГ), или кортикотропин, оказывает стимулирующее действие на кору надпочечников. В большей степени его влияние выражено на пучковую зону, что приводит к увеличению образования глюкокортикоидов, в меньшей - на клубочковую и сетчатую зоны, поэтому на продукцию минералокортикоидов и половых гормонов он не оказывает значительного воздействия. АКТГ усиливает синтез холестерина и скорость образования прегненолона из холестерина. Вненадпочечниковые эффекты АКТГ заключаются в стимуляции липолиза (мобилизует жиры из жировых депо и способствует окислению жиров), увеличении секреции инсулина и соматотропина, накоплении гликогена в клетках мышечной ткани, гипогликемии, что связано с повышенной секрецией инсулина, усилении пигментации за счет действия на пигментные клетки меланофоры. Продукция АКТГ подвержена суточной периодичности, что связано с ритмичностью выделения кортиколиберина. Максимальные концентрации АКТГ отмечаются утром в 6 - 8 часов, минимальные - с 18 до 23 часов. Образование АКТГ регулируется кортиколиберином гипоталамуса. Секреция АКТГ усиливается при стрессе, а также под влиянием факторов, вызывающих стрессогенные состояния: холод, боль, физические нагрузки, эмоции. Торможение продукции АКТГ происходит под влиянием самих глюкокортикоидов по механизму обратной связи. Избыток АКТГ приводит к гиперкортицизму, т.е. увеличенной продукции кортикостероидов, преимущественно глюкокортикоидов. Это заболевание развивается при аденоме гипофиза и носит название болезни Иценко-Кушинга. Основные проявления ее: гипертония, ожирение, имеющее локальный характер (лицо и туловище), гипергликемия, снижение иммунной защиты организма. Недостаток гормона ведет к уменьшению продукции глюкокортикоидов, что проявляется нарушением метаболизма и снижением устойчивости организма к различным влияниям среды. Тиреотропный гормон (ТТГ), или тиреотропин, активирует функцию щитовидной железы, вызывает гиперплазию ее железистой ткани, стимулирует выработку тироксина и трийодтиронина. Образование тиреотропина стимулируется тиреолиберином гипоталамуса, а угнетается соматостатином. Секреция тиреолиберина и тиреотропина регулируется йодсодержащими гормонами щитовидной железы по механизму обратной связи. Секреция тиреотропина усиливается также при охлаждении организма, что приводит к повышению выработки гормонов щитовидной железы и повышению тепла. Глюкокортикоиды тормозят продукцию тиреотропина. Секреция тиреотропина угнетается также при травме, боли, наркозе. Избыток тиреотропина проявляется гиперфункцией щитовидной железы, клинической картиной тиреотоксикоза. Фолликулостимулирующий гормон (ФСГ), или фоллитропин, вызывает рост и созревание фолликулов яичников и их подготовку к овуляции. У мужчин под влиянием ФСГ происходит образование сперматозоидов. Лютеинизирующий гормон (ЛГ), или лютропин, способствует разрыву оболочки созревшего фолликула, т.е. овуляции и образованию желтого тела. ЛГ стимулирует образование женских половых гормонов - эстрогенов. У мужчин этот гормон способствует образованию мужских половых гормонов - андрогенов. Секреция ФСГ и ЛС регулируется гонадолиберином гипоталамуса. Образование гонадолиберина, ФСГ и ЛГ зависит от уровня эстрогенов и андрогенов и регулируется по механизму обратной связи. Гормон аденогипофиза пролактин угнетает продукцию гонадотропных гормонов. Тормозное действие на выделение ЛГ оказывают глюкокортикоиды. Соматотропный гормон (СТГ), или соматотропин, или гормон роста, принимает участие в регуляции процессов роста и физического развития. Стимуляция процессов роста обусловлена способностью соматотропина усиливать образование белка в организме, повышать синтез РНК, усиливать транспорт аминокислот из крови в клетки. Наиболее ярко влияние гормона выражено на костную и хрящевую ткани. Действие соматотропина происходит посредством "соматомединов", которые образуются в печени под влиянием соматотропина. Обнаружено, что у пигмеев на фоне нормального содержания соматотропина не образуется соматомедин С, что, по мнению исследователей, служит причиной их маленького роста. Соматотропин влияет на углеводный обмен, оказывая инсулиноподобное действие. Гормон усиливает мобилизацию жира из депо и использование его в энергетическом обмене. Продукция соматотропина регулируется соматолиберином и соматостатином гипоталамуса. Если гиперфункция передней доли гипофиза проявляется в детском возрасте, то это приводит к усиленному пропорциональному росту в длину - гигантизму. Если гиперфункция возникает у взрослого человека, когда рост тела в целом уже завершен, наблюдается увеличение лишь тех частей тела, которые еще способны расти. Это пальцы рук и ног, кисти и стопы, нос и нижняя челюсть, язык, органы грудной и брюшной полостей. Это заболевание называется акромегалией. Причиной являются доброкачественные опухоли гипофиза. Гипофункция передней доли гипофиза в детстве выражается в задержке роста - карликовости ("гипофизарный нанизм"). Умственное развитие не нарушено. Пролактин стимулирует рост молочных желез и способствует образованию молока. Гормон стимулирует синтез белка - лактальбумина, жиров и углеводов молока. Пролактин стимулирует также образование желтого тела и выработку им прогестерона. Влияет на водно-солевой обмен организма, задерживая воду и натрий в организме, усиливает эффекты альдостерона и вазопрессина, повышает образование жира из углеводов. Образование пролактина регулируется пролактолиберином и пролактостатином гипоталамуса, а также в меньшей степени другими пептидами гипоталамуса . Секреция пролактина усиливается после родов и рефлекторно стимулируется при кормлении грудью. Эстрогены стимулируют синтез и секрецию пролактина. Избыток пролактина наблюдается при доброкачественной опухоли гипофиза, при менингитах, энцефалитах, травмах мозга, избытке эстрогенов, при применении некоторых противозачаточных средств. К его проявлениям относятся выделение молока у некормящих женщин (галакторея) и аменорея. Гормоны задней доли гипофиза Эти гормоны образуются в гипоталамусе. В нейрогипофизе происходит их накопление. В клетках супраоптического и паравентрикулярного ядер гипоталамуса осуществляется синтез окситоцина и антидиуретического гормона. Синтезированные гормоны путем аксонального транспорта с помощью белка - переносчика нейрофизина по гипоталамо-гипофизарному тракту - транспортируются в заднюю долю гипофиза. Здесь происходит депонирование гормонов и в дальнейшем выделение в кровь. Антидиуретический. гормон (АДГ), или вазопрессин, осуществляет в организме антидиуретическое действие, которое выражается в стимуляции реабсорбции воды в дистальном отделе нефрона. Это действие осуществляется благодаря взаимодействию гормона с вазопрессиновыми рецепторами типа V-2, что приводит к повышению проницаемости стенки канальцев и собирательных трубочек для воды, ее реабсорбции и концентрированию мочи. В клетках канальцев происходит также активация гиалуронидазы, что приводит к усилению деполимеризации гиалуроновой кислоты, в результате чего повышается реабсорбция воды и увеличивается объем циркулирующей жидкости. При недостаточности образования АДГ развивается несахарный диабет, или несахарное мочеизнурение, который проявляется выделением больших количеств мочи (до 25 л в сутки) низкой плотности, повышенной жаждой. Причинами несахарного диабета могут быть острые и хронические инфекции, при которых поражается гипоталамус (грипп, корь, малярия), черепно-мозговые травмы, опухоль гипоталамуса. Избыточная секреция АДГ ведет, напротив, к задержке воды в организме. Окситоцин избирательно действует на гладкую мускулатуру матки, вызывая ее сокращения при родах. На поверхностной мембране клеток существуют специальные окситоциновые рецепторы. Во время беременности окситоцин не повышает сократительную активность матки, но перед родами под влиянием высоких концентраций эстрогенов резко возрастает чувствительность матки к окситоцину. Окситоцин участвует в процессе лактации. Усиливая сокращения миоэпителиальных клеток в молочных железах, он способствует выделению молока. Увеличение секреции окситоцина происходит под влиянием импульсов от рецепторов шейки матки, а также механорецепторов сосков грудной железы при кормлении грудью. Эстрогены усиливают секрецию окситоцина. Функции окситоцина в мужском организме изучены не достаточно. Недостаток продукции окситоцина вызывает слабость родовой деятельности. Щитовидная железа Щитовидная железа состоит из двух долей, соединенных перешейком и расположенных на шее по обеим сторонам трахеи ниже щитовидного хряща. Она имеет дольчатое строение. Ткань железы состоит из фолликулов, заполненных коллоидом, в котором имеются йодсодержащие гормоны тироксин (тетрайодтиронин) и трийодтиронин в связанном состоянии с белком тиреоглобулином. В межфолликулярном пространстве расположены парафолликулярные клетки, которые вырабатывают гормон тиреокальцитонин. Содержание тироксина в крови больше, чем трийодтиронина. Однако активность трийодтиронина выше, чем тироксина. Эти гормоны образуются из аминокислоты тирозина путем ее йодирования. Инактивация происходит в печени посредством образования парных соединений с глюкуроновой кислотой. Йодсодержащие гормоны выполняют в организме следующие функции: 1) усиление всех видов обмена (белкового, липидного, углеводного), повышение основного обмена и усиление энергообразования в организме; 2) влияние на процессы роста, физическое и умственное развитие; 3) увеличение частоты сердечных сокращений; 4) стимуляция деятельности пищеварительного тракта: повышение аппетита, усиление перистальтики кишечника, увеличение секреции пищеварительных соков; 5) повышение температуры тела за счет усиления теплопродукции; 6) повышение возбудимости симпатической нервной системы. Секреция гормонов щитовидной железы регулируется тиреотропным гормоном аденогипофиза, тиреолиберином гипоталамуса, содержанием йода в крови. При недостатке йода в крови, а также йодсодержащих гормонов по механизму положительной обратной связи усиливается выработка тиреолиберина, который стимулирует синтез тиреотропного гормона, что, в свою очередь, приводит к увеличению продукции гормонов щитовидной железы. При избыточном количестве йода в крови и гормонов щитовидной железы работает механизм отрицательной обратной связи. Нарушения функции щитовидной железы проявляются ее гипофункцией и гиперфункцией. Если недостаточность функции развивается в детском возрасте, то это приводит к задержке роста, нарушению пропорций тела, полового и умственного развития. Такое патологическое состояние называется кретинизмом. У взрослых гипофункция щитовидной железы приводит к развитию патологического состояния - микседемы. При этом заболевании наблюдается торможение нервно-психической активности, что проявляется в вялости, сонливости, апатии, снижении интеллекта, нарушении половых функций, угнетении всех видов обмена веществ и снижении основного обмена. У таких больных увеличена масса тела за счет повышения количества тканевой жидкости и отмечается одутловатость лица. Отсюда и название этого заболевания: микседема - слизистый отек Гипофункция щитовидной железы может развиться у людей проживающих в местностях, где в воде и почве отмечается недостаток йода. Это так называемый эндемический зоб. Щитовидная железа при этом заболевании увеличена (зоб), возрастает количество фолликулов, однако из-за недостатка йода гормонов o6разуется мало, что приводит к соответствующим нарушениям в организме, проявляющимся в виде гипотиреоза. При гиперфункции щитовидной железы развивается заболевание тиреотоксикоз (диффузный токсический зоб, Базедова болезнь, болезнь Грейвса). Характерными признаками этого заболевания являются увеличение щитовидной железы (зоб) экзофтальм, тахикардия, повышение обмена веществ, особенно основного, потеря массы тела, увеличение аппетита, нарушение теплового баланса организма, повышение возбудимости и раздражительности. |