Контрольная работа. рыба. И осморегуляция. 3
 Скачать 1.86 Mb. Скачать 1.86 Mb.
|

|
Содержание 1.Выделительная система и осморегуляция. 3 2.Различие в строении осевого скелета и черепа у рыбообразных (круглоротых), хрящевых (акул, скатов) и костистых рыб. 9 3.Реакция рыб на электросвет, электроток и ультразвук. 15 4Тресковые рыбы северных морей и морей Дальнего Востока. 17 Общие признаки и различия 18 Треска атлантическая 19 5 Отличия в симптомах при злокачественной и незлокачественной краснухе карпов. Когда наблюдается наибольшее заболевание карпов краснухой? 23 6Ушастые тюлени (сивучи, морские котики). Особенности строения, черты биологии, распространение, полезная продукция. 24 8Важнейшие промысловые рыбы Аральского моря. Динамика их уловов. 28 9Место рыб в водных биоценозах. 29 10 Методика составления промысловых карт. 34 Список литературы 36 Выделительная система и осморегуляция.Ответ. В отличие от высших позвоночных, имеющих компактную тазовую почку (метанефрос), рыбы обладают более примитивной туловищной почкой (мезонефрос), а их зародыши - предпочкой (пронефрос). У некоторых видов (бычок, атерина, бельдюга, кефаль) предпочка в том или ином виде выполняет выделительную функцию и у взрослых особей. У большинства же взрослых рыб функционирующей почкой становится мезонефрос. Почки - парные, вытянутые вдоль полости тела тёмно-красные образования, плотно прилегающие к позвоночнику над плавательным пузырём (рис. 24). В почке выделяют передний (головная почка), средний и задний отделы, артериальная кровь поступает в почки по почечным артериям, венозная - по воротным венам почек. Морфофизиологическим элементом почки является извитой почечный мочевой каналец, один конец которого расширяется в мальпигиево тельце, а другой отходит к мочеточнику. Мальпигиево тельце - клубочек артериальных капилляров (гломерула), охватываемый расширенными стенками канальца, - образует боуменову капсулу. У примитивных форм (акулы, скаты, осетровые) перед капсулой от канальца отходит мерцательная воронка. Мальпигиев клубочек служит аппаратом для фильтрации жидких продуктов обмена. В фильтрат попадают как продукты обмена, так и важные для организма вещества. Стенки почечных канальцев пронизаны капиллярами воротных вен и сосудов из боуменовых капсул. Железистые клетки стенок секретируют продукты азотистого распада (мочевину), которые попадают в просвет канальцев. Здесь же, в стенках канальцев, происходит обратное всасывание воды, сахаров, витаминов из фильтрата мальпигиевых телец. Очищенная кровь возвращается в сосудистую систему почек (почечную вену), а отфильтрованные из крови продукты обмена и мочевина выводятся через каналец и мочеточник. Мочеточники изливаются в мочевой пузырь (мочевой синус), а затем моча выводится наружу. У самцов большинства костистых рыб через мочеполовое отверстие позади ануса, а у самок костистых и самцов лососевых, сельдей, щук и некоторых других -через анальное отверстие. У акул и скатов мочеточник открывается в клоаку. Передний отдел почки - головная почка выполняет не выделительную, а кроветворную функцию: в него не заходит воротная вена почек, а в составляющей её лимфоидной ткани образуются красные и белые кровяные клетки и разрушаются отжившие эритроциты. Как и селезёнка, почки чутко отражают состояние рыбы, уменьшаясь в объёме при недостатке кислорода в воде и увеличиваясь при замедлении обмена (у карпа - во время зимовки, когда ослабляется деятельность кровеносной системы), в случае острых заболеваний и др. Очень своеобразна дополнительная функция почек у колюшки, строящей для нереста гнездо из кусочков растений: перед нерестом почки увеличиваются, в стенках почечных канальцев вырабатывается большое количество слизи, которая в воде быстро затвердевает и скрепляет гнездо. Строение почки и схема почечных канальцев рыб приведены на рисунке 1.  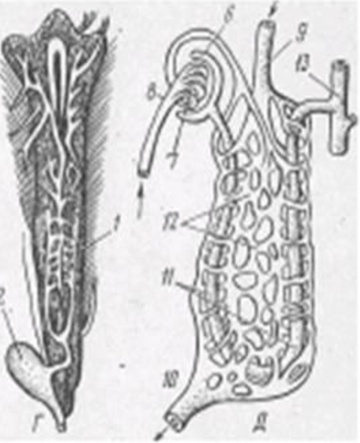 Рисунок 1 - Почка форели и схема почечных канальцев рыб А — головная почка (пред- почка); Б, В, Д— туловищная почка; Г — почка форели; 1 — мочеточники; 2 — мочевой пузырь; 3 — наружный клубочек; 4 — мальпигиево тельце; 5 — воронка; 6 — капсула; 7 — сосудистый клубочек; 8 — ветвь спинной аорты; 9 — воротная вена почек; 10 — почечная вена; 11 — почечные канальцы; 12 — венозные синусоиды; 13 — собирательная трубка. В процессах выделения и водно-солевого обмена кроме почек принимают участие кожа, жаберный эпителий, пищеварительная система. Вода - существенный компонент живого организма. Будучи универсальным растворителем, является средой, обеспечивающей ход биологических процессов на клеточном уровне. Жизненная среда рыб - морские и пресные воды - всегда имеют большее или меньшее количество солей, поэтому осморегуляция является важнейшим условием жизнедеятельности, а осморегуляторные приспособления - специфическим для рыб. Солевой состав воды также создает определенные проблемы для рыб. Содержание воды в организме рыб составляет 60-80%, содержание минеральных веществ -0,6-1%. У морских рыб и миксин солевой состав крови практически не отличается от солевого состава среды их обитания. Поэтому у них нет серьезного осмотического барьера. И в пресной воде морские виды очень быстро погибают из-за нарушения водно-солевого состава тела. У пресноводных и проходных рыб механизм осморегуляции очень эффективен и заслуживает отдельного рассмотрения. Все виды животных имеют механизм осморегуляции. Осмотическое давление клетки - это основа мембранного потенциала, определяющего важнейшие свойства тканей - раздражимость и возбудимость. У рыб можно выделить два основных механизма осморегуляции. Первый (примитивный) механизм осморегуляции характерен для акул. Плазма, лимфа и другие жидкости организма этих рыб по электролитному составу мало отличаются от морской воды, соленость которой равнозначна 1% NaCl. На первый взгляд в таких условиях отпадает необходимость тратить энергию на осморегуляцию. Однако специфика азотистого обмена такова, что в крови акул накапливается большое количество (5%) мочевины и триметиламиноксида, которые плохо диффундируют через мембраны клеток почек и жабр. Поэтому у акул возникает повышенное осмотическое давление внутренней среды (26атм против 24атм в морской воде) (1атм= 105Па). Это создает условия для свободного проникновения воды из внешней среды в межклеточную жидкость и кровь, что, в свою очередь, меняет градиент концентрации NaCl и приводит к диффузии натрия и калия. Фактически, мембраны удерживают только мочевину. Организм акулы представляет собой устойчивую биологическую систему, т. е. обладает гомеостазом, как и у всех других животных. Другими словами, у акулы есть механизм поддержания электролитного состава. Эволюция снабдила акул для поддержания гомеостаза специальным органом ректальной железой, которая выводит в клоаку избыток солей. Второй тип осморегуляции присущ костистым рыбам. Этот механизм более эффективен, однако и более сложен. Костистые обитают как в пресной, так и в соленой воде. Поэтому и внутри этой группы есть некоторые различия в механизме осморегуляции. У пресноводных костистых рыб осмоляльность крови выше, чем окружающей их воды: соответственно 6 и 0,3 атм. В соленой воде картина обратная: осмотическое давление крови составляет 9 атм, а морской воды - 24 атм. Следовательно, и в первом, и во втором случае требуются затраты энергии для поддержания осмотического давления внутренней среды организма рыб. Осмотическое давление водных животных создаётся давлением их полостных жидкостей, давлением крови и соков тела. Определяющая роль в этом процессе принадлежит водно-солевому обмену. Каждая клетка тела имеет оболочку: она полупроницаема, то есть по-разному проницаема для воды и солей (пропускает воду и соли избирательно). Водносолевой обмен клеток определяется, в первую очередь осмотическим давлением крови и клеток. По уровню осмотического давления внутренней среды по отношению к окружающей воде рыбы образуют несколько групп: 1) у миксин полостные жидкости изотоничны окружающей среде; 2) у акул и скатов концентрация солей в жидкостях тела и осмотическое давление немного выше, чем в морской воде, или почти равно ему (достигается за счёт разницы солевого состава крови и морской воды, и за счёт мочевины); 3) у костистых рыб, как морских, так и пресноводных (как и у более высокоорганизованных позвоночных), осмотическое давление внутри тела не равно осмотическому давлению окружающей воды. У пресноводных оно выше, чем у морских рыб (как и у других позвоночных). У пресноводных рыб осмотическое давление эквивалентно воде солёностью 6 - 7%о, у морских костистых - 7 - 8%о, у акул - до 30%о. Если в организме поддерживается определённый уровень осмотического давления жидкостей тела, то условия жизнедеятельности клеток становятся более стабильными и организм меньше зависит от колебаний внешней среды. Настоящие рыбы способны сохранять постоянное осмотическое давление крови и лимфы, то есть внутренней среды, поэтому их относят к гомойосмотическим организмам (от греч. «гомойос» -однородный). Но у разных групп рыб эта независимость осмотического давления выражается и достигается по-разному: 1) у морских костистых рыб общее количество солей в крови ниже, чем в морской воде, давление внутренней среды меньше давления внешней, то есть их кровь гипотонична по отношению к морской воде; 2) у пресноводных рыб количество солей в крови выше, чем в пресной воде. Давление внутренней среды больше давления внешней, их кровь гипертонична. Поддержание солевого состава крови и давления на нужном уровне обусловливается деятельностью: почек, особых клеток стенок почечных канальцев (выделение мочевины), жаберных лепестков (диффузия аммиака, выделение хлоридов), кожных покровов, кишечника, печени. У морских и пресноводных рыб осморегуляция совершается разными способами: специфическая деятельность почек, различная проницаемость покровов для мочевины, солей и воды, различная деятельность жабр в морской и пресной воде. У пресноводных рыб (с гипертонической кровью), находящихся в гипотонической среде, разница осмотического давления внутри и вне организма приводит к тому, что вода извне непрерывно поступает внутрь организма - через жабры, кожу и ротовую полость (рис. 2). 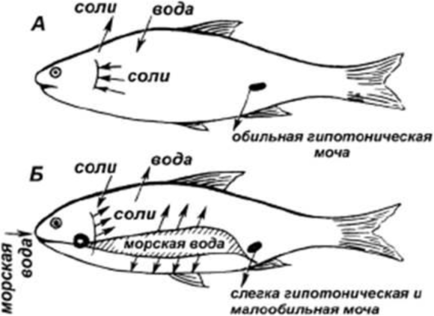 Рисунок 2 - Основные пути движения воды и солей в процессе осморегуляции у пресноводных и морских костистых рыб: А— пресноводные; Б — морские костистые рыбы При этом основной приток осмотической воды проходит через легко пропускающий воду жаберный эпителий (около 1 км3 воды в сутки). Утрата солей с мочой, экскрементами и через кожу восполняется у пресноводных рыб за счёт получения их с пищей благодаря специализированной деятельности жабр (жабры поглощают из пресной воды ионы Na и С1) и поглощению солей в почечных канальцах. Морские костистые рыбы (с гипотонической кровью), находящиеся в Процессы осморегуляции регулируются нейросекреторной системой: аденогипофиз, урогипофиз, интерреналовая ткань, щитовидная железа и др. В морской воде покровные ткани костистых рыб избирательно пропускают воду и не пропускают растворенные в ней соли. Рыбы активно заглатывают воду, однако слизистая желудочно-кишечного тракта (ЖКТ) избирательно абсорбирует только воду и одновалентные ионы натрия, калия, хлора. Остальные удаляются из организма в составе каловых масс. Вероятный избыток двухвалентных ионов (кальций, магний, сульфат) выводится почками. Помимо этого имеется и дополнительный механизм регуляции ионного состава. Слизистая жаберного аппарата включает в себя сеть специфических клеток, которые экскретируют во внешнюю среду ионы хлора и натрия. Они так и называются - "хлоридные". Необходимо подчеркнуть, что жабры имеют особый мембранный потенциал, который облегчает экскрецию катионов. Этот потенциал оценивается в 20-25 мВ. Учитывая, что снаружи жаберная мембрана несет положительный заряд, экскреция натрия в таких условиях становится довольно эффективной даже без затрат энергии. В пресной воде угрозу гомеостазу создает сама вода (не электролиты). Поэтому заглатывание воды пресноводными рыбами биологически нецелесообразно. Однако через жабры вода диффундирует активно. Компенсируется эта избыточность очень эффективной работой почек. У костистых рыб в пресной воде образуются и выделяются сравнительно большие количества мочи. По крайней мере, эта величина на порядок выше, чем у морских пластинчатожаберных. Почки пресноводных имеют очень высокую эффективность реабсорбции электролитов. Солевой гомеостаз регулируется нейрогуморальным путем. Осморецепторы располагаются не только в кровеносных сосудах, но и на слизистых жабр и ротовой полости. Их возбуждение приводит к активизации гипоталамо-гипофизарного механизма. Непосредственно на проницаемость мембран, через которые осуществляется транспортирование электролитов и воды, влияют соматотропин, пролактин, вазопрессин, окситоцин, кортизол. Помимо проницаемости мембран эти гормоны контролируют синтез белков - переносчиков ионов, влияют на регуляцию кровотока, фильтрации и реабсорбции в почках. Жаберные мембраны более активно пропускают электролиты за счет активного переноса и диффузии по сравнению с водой (только диффузия). Мембранный потенциал жаберных мембран костистых рыб в пресной воде составляет минус 30 - минус 40 мВ, что благоприятствует переносу катионов из внешней среды в кровь. |