Основы физиологии почки. Реферат. Реферат Почка. И. П. Павлова Федерального агентства но здравоохранению и социальному развитию Кафедра нормальной физиологии Студент Факультет Основы физиологии почки Реферат
 Скачать 0.55 Mb. Скачать 0.55 Mb.
|

|
концентрационному показателю (Ux/Px), то есть отношению концентрации вещества в конечной моче (Ux) к концентрации в крови (Рх). Такое отношение концентраций по инулину (Пин/Рин) указывает, во сколько раз уменьшился объем фильтрата при прохождении по канальцам. Если концентрационный показатель для какого-либо вещества больше показателя по инулину, то это вещество в почках секретируется, а если меньше - то реабсорбируется. Отношение концентрационного показателя вещества к концентрационному показателю по инулину называется экскретируемой фракцией (EF). СЕКРЕЦИЯ ИОНОВ ВОДОРОДА Секреция ионов водорода происходит в проксимальном канальце, толстой восходящей части петли Генле, в системе собирательной трубки и играет важную роль в регуляции pH крови. От секреции иона водорода зависит реабсорбция НСО3. Реабсорбция в кровь одного иона НСО3 оказывает такой же эффект на pH крови, как и удаление иона водорода из нее. В процессе реабсорбции бикарбонатов важную роль играет обмен части реабсорбируемых ионов Na+ на внутриклеточные ионы водорода, которые механизмом антипорта переходят в просвет канальцев. В канальцевой жидкости ионы Н+, соединяясь с ионами НСО3, образуют угольную кислоту, которая при содействии карбоангидразы щеточной каймы клеток канальцев расщепляется на СО2 и Н2О. Растворенный СО2 легко проникает путем диффузии в клетки эпителия канальцев, где взаимодействует с водой. В результате этой реакции образуется снова угольная кислота, которая внутри клеток канальцев при участии карбоангидразы диссоциирует на ионы Н+ и НСО3. Водород активно секретируется в просвет канальца, а НСО3 реабсорбируется через базолатсральную мембрану в интерстициальную жидкость путем облегченной диффузии и поступает в кровь перитубулярных капилляров. Посредством секреции ионов ЕГ в проксимальном канальце реабсорбируется примерно 80% профильтровавшегося бикарбоната. В толстой восходящей части петли Генле реабсорбируется 10-15% бикарбоната, а почти весь оставшийся бикарбонат - в дистальном извитом канальце и системе собирательной трубки. Кроме того, секретируемый ион водорода в просвете канальца соединяется с небикарбонатным буфером, в основном с фосфатом (НРО4 ), образуется Н2РО4. Таким путем ионы водорода, связанные с фосфатным буфером, выводятся с мочой. Для перемещения через мембрану как иона водорода, так и бикарбоната, необходимы специальные переносчики. Активный перенос иона водорода через апикальную мембрану из клетки в просвет канальца осуществляется тремя различными переносчиками. Первично активная Н+-АТФаза присутствует во всех секретирующих ион водорода сегментах канальца и собирательной трубке. В проксимальном канальце и толстой восходящей части петли Генле имеется большое количество Na+-H+ переносчиков - антипортеров, поэтому секреция ионов Н+ в этих сегментах, в основном, осуществляется вторично активным транспортом и стимулируется реабсорбцией иона Na+. В собирательной трубке имеются вставочные клетки типа А. На их мембране локализованы К+,Н+-АТФазы, которые одновременно переносят ионы водорода в просвет канальца, а ионы К+ в клетку. Еще один механизм регулирования кислотно-щелочного баланса реализуется почечной глутаминазой. Под ее влиянием осуществляется гидролиз глутамина на глутамат и аммиак. Аммиак секретируется в просвет канальца, где соединяется с ионом Н+ и выводится с конечной мочой в виде ионов аммония. ПОСТУПЛЕНИЕ ЖИДКОСТИ В ПЕРИТУБУЛЯРНЫЕ КАПИЛЛЯРЫ Перенос веществ из интерстициального пространства в кровь перитубулярных капилляров (рис. 9) обусловлен тем, что силы, определяющие реабсорбцию (Рр) тканевой жидкости в кровь, больше сил, способствующих фильтрации (Рф) из крови в иитерстиций. Реабсорбцию обеспечивает сумма гидростатического давления жидкости интерстиция (Рги= 3 мм рт. ст.) и онкотического давление крови (Рок= 33 мм рт.ст.), а фильтрацию - сумма гидростатического давления крови (Ргк= 20 мм рт.ст.) и онкотического давления в интсрстиции (Рои= 6 мм рт.ст.): Рр = Рги + Рок = 36 мм рт.ст. Рф = Ргк + Рои = 26 мм рт.ст. 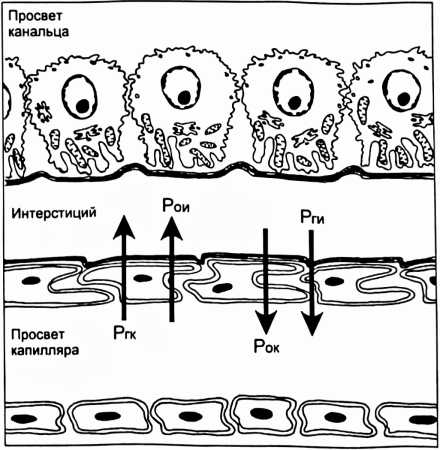 Рис. 9. Транспорт ионов и молекул из канальцевой жидкости в кровь перитубулярных капилляров. Стрелками показаны силы, способствующие (■!•) или препятствующие (Т) транспорту веществ из интерстиция в кровь перитубулярных капилляров. МЕХАНИЗМ СОЗДАНИЯ КОНЦЕНТРАЦИОННЫХ ГРАДИЕНТОВ В ПОЧКЕ Конечная моча может быть как гипотонической, так и гипертонической, особенно при повышении осмотического давления крови, когда необходимо экономить воду и удалять из внутренней среды организма осмотически активные вещества. Способностью выделять гипертоническую мочу обладают только почки млекопитающих и птиц. Механизм концентрирования мочи обусловлен деятельностью поворотно-противоточной системы. Поворотно-противоточная система представлена петлей Генле, собирательной трубкой, прямыми кровеносными сосудами (vasa recta) и ин- терстицием мозгогового вещества почки. Главный элемент системы - петля Генле состоит из нисходящего тонкого колена, которое у вершины почечного сосочка поворачивается под углом 180° и переходит в восходящее колено, имеющее вначале тонкий, а затем толстый отдел, который еще называют прямой частью дистального канальца. В нисходящее колено поступает за сутки около 60 л изотоничной жидкости из проксимального извитого канальца. Стенка нисходящего колена хорошо проницаема для воды и почти не пропускает ионы СГ и Na+. Стенка восходящего колена не проницаема для воды, но через нее активно транспортируются ионы С1 и Na+ в окружающий интерстиций, повышая его осмотическое давление (рис. 10). По осмотическому градиенту вода выходит из нисходящего колена петли в интерстиций. Жидкость в нисходящем колене становится гипертонической, причем ее осмотическое давление равным давлению в интерстиции на данном уровне мозгового вещества почки. Несмотря на поступление воды из нисходящего колена, осмотическое давление в интерстиции не снижается из-за продолжающегося активного транспорта ионов С1 и Na+ из восходящего колена. Эти процессы совершаются на всем пути движения жидкости по петле, поэтому происходит постепенное увеличение осмотичности в нисходящем колене и в интерстиции по мере удаления от корковой зоны вглубь мозгового вещества. Во время движения жидкости по восходящему колену ее осмотич- ность понижается из-за активного транспорта ионов, и она становится гипотонической. Возникает поперечный осмотический градиент. В корковую зону из восходящего колена притекает около 20 л гипотоничной жидкости в сутки. собирательной трубки мочевина выходит в иитерстиций, попадает в кровь и в жидкость, текущую по петле Генле. Осмотичность названных сред возрастает, что способствует увеличению реабсорбции воды. Из петли Генле мочевина попадает в собирательную трубку, из нее в иитерстиций и т.д., наблюдается кругооборот мочевины. При движении крови по vasa recta в глубь мозгового вещества в нее поступают ионы Na+, Cl и пассивно выходит вода. При движении крови из мозгового вещества в корковое в нее пассивно поступает вода и выходят ионы Na+, Cl. Таким образом, vasa recta функционируют как противоточный обменник, который препятствует вымыванию из интерстиция осмотически активных ионов. Повышение кровотока в прямых сосудах ведет к вымыванию осмотически активных веществ и уменьшению реабсорбции воды. Какая-то часть воды с растворенными в нем веществами из интерстиция попадает в кровь перитубулярных капилляров и уносится из почки. Благодаря поворотно-противоточной системе, реабсорбируется около 60 л воды в сутки, почти весь натрий, не успевший реабсорбироваться в проксимальных канальцах. Механизм большинства диуретиков базируется на их способности уменьшать реабсорбцию ионов, прежде всего Na+, что ведет к снижению реабсорбции воды и увеличению диуреза. РЕНИН-АНГИОТЕНЗИН-АЛЬДОСТЕРОНОВАЯ СИСТЕМА Большое значение в регуляции артериального давления, почечного кровотока и работы почки имеет ренин-ангиотензин-альдостероновая система (РААС). В месте соприкосновения дистального извитого канальца с сосудами: vas afference и vas efference располагается юкстагломерулярный аппарат почки (ЮГА), имеющий форму треугольника (рис.11). Основание этого треугольника образовано гиперхромными клетками канальца - плотным пятном (macula densa). Боковые стороны образованы гранулированными эпителиальными - юкстагломерулярными клетками, лежащими в артериолах, особенно их много в vas afference. Основная функция этих клеток - секреция ренина. Клетки macula densa играют роль своеобразных рецепторов, реагирующих на изменение концентрации ионов Na+ и Cl в жидкости, протекающей в дистальном канальце. При увеличении почечного кровотока растет объем ультрафильтрата и скорость его движения по канальцам. В связи с этим в проксимальных канальцах не успевает пеявелпПкплпатыа плпжняя часть профильтровав- шихся ионов Na+ и С1. Поэтому увеличивается загрузка ими дистальных отделов канальцев, а также реабсорбция их клетками macula densa. 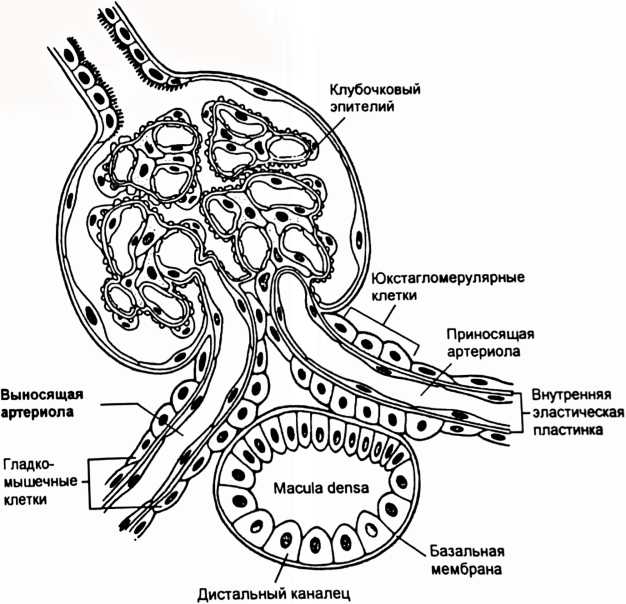 Рис. 11. Структура юкстагломерулярного аппарата (Guyton А.С., Hall Y.E., 2001). При увеличении концентрации ионов Na+ и Cl в просвете канальца, Na+, К+, С1 переносчик на апикальной мембране клеток плотного пятна способствует повышению содержания этих ионов и в цитоплазме эпителиальных клеток. В результате открытия катионных каналов плазмолеммы происходит поступления Са++ в цитоплазму. Прирост Са** в цитоплазме стимулирует секрецию из клеток плотного пятна паракринных агентов в виде аденозина. Гладкомышечные клетки приносящей артериолы имеют рецепторы к аденозину. Воздействие аденозина на эти рецепторы приводит к поступлению в цитоплазму мышечных клеток ионов Са++, а вызванная ими кон- стрикция артериолы снижает СКФ. При уменьшении общего объема жидкости в организме снижается почечный кровоток и скорость клубочковой фильтрации, и поэтому в дистальный каналец притекает меньше жидкости с ионами натрия и хлора. Снижение концентрации в дистальных канальцах ионов Na+ и CI улавливается клетками macula densa, последние посылают сигнал к юкстагломерулярным клеткам, что стимулирует секрецию ренина (рис. 12). Такая система сигнал - эффектор позволяет почкам регулировать реабсорбцию ионов Na+ и осуществлять контроль над АД, почечным кровотоком, электролитным составом крови. Ренин является ферментом, который действует на белок крови ангиотензиноген (а-глобулин), который вырабатывается в печени и циркулирует в крови. Ангиотензиноген в присутствии ренина превращается в ангиотензин I, который под влиянием ангиотензинпревращающего фермента переходит в ангиотензин II. Ангиотензин II вызывает разнообразные биологические эффекты: сокращение гладких мышц кровеносных сосудов и мезангиоцитов, стимулирует жажду и секрецию альдостерона, но тормозит секрецию ренина, являясь одним из звеньев отрицательной обратной связи в регуляции секреции ренина. Растяжение гранулярных клеток при увеличении почечного кровотока и давления в приносящей артериоле угнетает секрецию ренина. Большинство этих эффектов ведет к поддержанию уровня кровяного давления. В клинике часто для снижения артериального давления используются препараты, ингибирующие ангиотензинпревращающий фермент. Секреция ренина усиливается через Ргадренорецепторы симпатической нервной системой, а также под влиянием адреналина, АДГ, при повышении концентрации К* и снижении Na+ в крови. Секреция ренина снижается под влиянием предсердного натрийуретического гормона, ангиотензина II, аденозина. Ангиотензин II стимулирует секрецию альдостерона корой надпочечников. Альдостерон проходит через базолатеральную мембрану клеток почечных канальцев и соединяется с рецепторами в цитоплазме. Гормонрецеп- торный комплекс проникает в ядро клетки и влияет на синтез РНК, что ведет к образованию, как минимум, двух белков. Один из них активирует 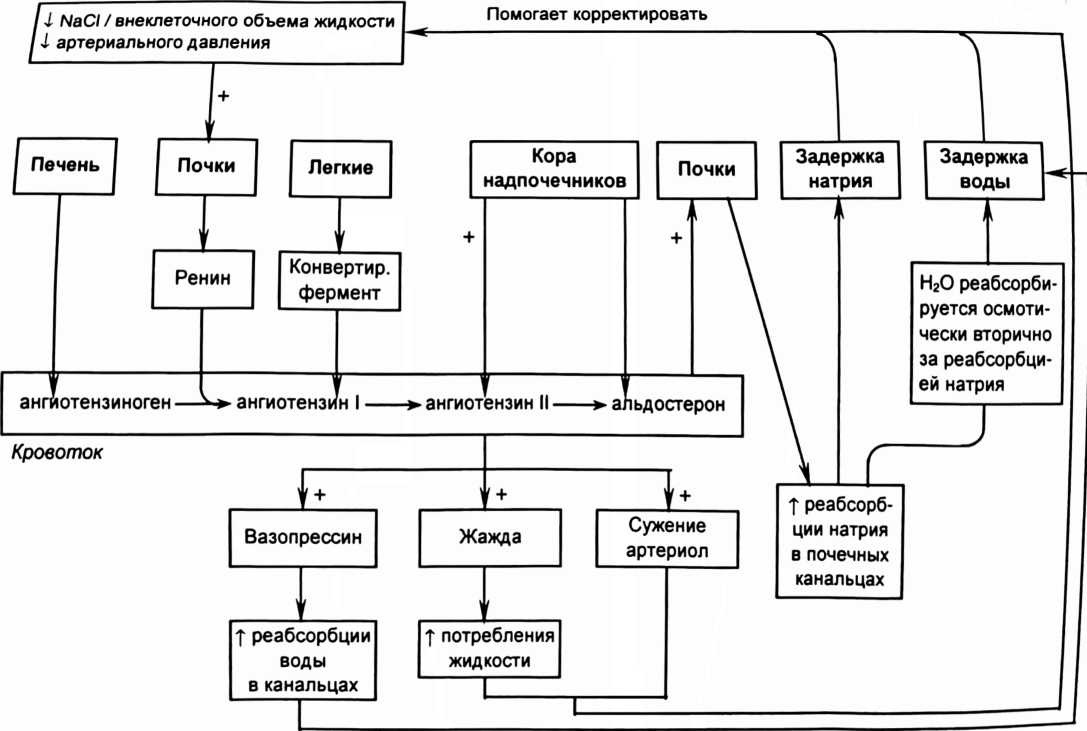 to Рис. 12. Ренин-ангиотензин-альдостероновая система (Sherwood L.. 1999). Na'.K -АТФазу на базолатеральной мембране, другой увеличивает проницаемость апикальной мембраны для иона Na*. НЕЙРОГУМОРАЛЬНЫЕ МЕХАНИЗМ Ы РЕГУЛЯЦИИ РАБОТЫ ПОЧКИ К почкам подходит большое количество симпатических норадренергических нервов, иннервирующих гладкую мускулатуру сосудов почки, юкстагломерулярный аппарат и многие участки канальца. Через Об-адренорецепторы симпатические нервы вызывают сосудосуживающий эффект в почке. В покое влияние симпатических нервов на сосуды почки мало и они не оказывают существенного влияния на почечную гемодинамику. Повышение тонуса симпатических нервов вызывает спазм артериол в почке и снижение почечного кровотока. Однако сужение артериол не обязательно вызовет снижение скорости клубочковой фильтрации (СКФ), поскольку констрикция обеих артериол ведет к незначительному повышению фильтрационного давления. Кроме того, уменьшение почечного кровотока стимулирует секрецию простагландина Е в почке. Простагландин расширяет сосуды почки, кровоток несколько возрастает, а в результате СКФ практически остается неизменной. Сосудистые эффекты симпатической регуляции связаны также с уменьшением кровотока в сосудах мозгового вещества почки, от которого зависит эффективность противоточной системы и концентрирования мочи. Симпатические нервы через [3-адренорецепторы стимулируют выделение ренина юкстагломерулярными клетками. Активация ренин-ангио- тензин-альдостероновой системы ведет к повышению артериального давления и увеличению реабсорбции натрия. Перерезка симпатических нервов, иннервирующих почки, ведет к повышению выведения ионов Na+ с конечной мочой (натрийурезу). Симпатические нервы оказывают трофические влияния на канальцы, повышая их реабсорбционную способность. Влияние парасимпатических нервов на почку выражено слабо. Стимуляция парасимпатических нервов снижает реабсорбцию натрия и увеличивает реабсорбцию глюкозы. Гуморальные влияния на почку выражены сильнее, чем нервные. Большое значение в регуляции почечного кровотока и работы почки имеет |