лекция. Катаболизм. Химическое строение и биологическая роль липидов. окисление жирных кислот и окисление глицерола. Липолиз
 Скачать 411.8 Kb. Скачать 411.8 Kb.
|

|
Катаболизм. Химическое строение и биологическая роль липидов. β-окисление жирных кислот и окисление глицерола. Липолиз - то расщепление ТАГ до глицерина и жирных кислот. Катализируется триглицеридлипазой, диглицеридлипазой и моноглицеридлипазой, которые действуют соответственно на ТАГ, ДАГ и МАГ. Триглицеридлипаза является лимитирующим ферментом липолиза. Она имеет две формы: фосфорилированную (активную) и нефосфорилированную (неактивную). Превращение неактивной формы в активную происходит под влиянием протеинкиназы. Протеинкиназа, в свою очередь, активируется в результате присоединения к ее аллостерическим центрам цАМФ. Регуляция липолиза происходит под действием гормонов. Активируют липолиз: катехоламины, глюкагон, СТГ, АКТГ, тироксин, липотропин гипофиза, цАМФ. Ингибирует липолиз инсулин.   МЕТАБОЛИЗМ ВЖК ВЖК из жировой ткани поступают в кровь и с помощью альмуминов транспортируются в различные органы и ткани. Пути поступления и использования ВЖК. Липиды пищи Липолиз в жировой ткани Синтез de novo Синтез ТАГ в жировой ткани Синтез сложных липидов Синтез кетоновых тел Синтез стероидов Окисление до СО2 и воды Активация жирной кислоты. Активация жирной кислоты происходит в цитозоле под действием ферментов ацил-КоА-синтетаз (тиокиназ). Известен ряд тиокиназ, специфичных к жирным кислотам с разной длиной углеводородной цепи. Эти ферменты в клетках прокариот прикреплены к клеточной мембране. Реакция протекает в 2 этапа и требует АТФ, HS-КоА и Mg2+. Вначале жирная кислота взаимодействует в АТФ с образованием ациладенилата, который затем реагирует с HS-КоА. Для активации используется энергия 2-х макроэргических связей, так как АТФ превращается в пирофосфат, который гидролизуется пирофосфатазой до неорганического фосфата.  Транспорт в митохондрии- трехэтапный перенос: Образованный ацил-КоА переносится через мембрану митохондрий с помощью карнитина (-гидрокси--триметиламиномасляная кислота) и ферментов, локализованных в цитозоле и митохондриях - карнитинацил-трансфераз. В цитозоле под действием карнитинацилтрансферазы I образуется ацил-карнитин, который транспортируется через мембрану митохондрий облегченной диффузией с использованием ацил-карнитин/карнитинового переносчика. В митохондриях происходит обратный процесс: под действием митохондриального фермента карнитин-ацилтрансферазы II (локализован на внутренней стороне внутренней мембраны митохондрий) ацилкарнитин взаимодействует с HS-КоA и освобождается ацил-КоА.  Процесс окисления условно делят на 2 этапа: Процесс окисления условно делят на 2 этапа: 1) процесс β-окисления; 2) окисление ацетил-КоА в ЦТК и сопряженных цепях переноса электронов. Окислительное расщепление жирных кислот – универсальный процесс, протекающий во всех видах живых организмов. 1. У млекопитающих окисление жирных кислот с энергетической целью происходит в печени, почках, скелетной и сердечной мышцах. В мозге, эритроцитах и мозговом слое надпочечников жирные кислоты не окисляются 2. Ферменты окисления жирных кислот локализованы в матриксе митохондрий. β-Окисление жирной кислоты. Первые предположения о путях окисления жирных кислот высказал Ф.Кнооп еще в 1904 г., выдвинув свою гипотезу «β-окисления»- цикл Кноопа- Линена, в соответствии с которой происходит последовательное отщепление двухуглеродных фрагментов с карбоксильного конца молекулы. С16→С14→С12→С10→С8→С6→С4→2С2 β-Окислению подвергаются только ВК с четным числом атомов С. В результате каждого цикла β-окисления, включающего 4 реакции, происходит высвобождение ацетил-КоА и жирная кислота укорачивается на 2 углеродных атома. 1. Дегидрирование. Ацил-КоА подвергается дегидрированию под действием фермента ацил-КоА-дегидрогеназы, содержащей в качестве простетической группы ФАД. Образуется двойная связь между α и β углеродными атомами. Фермент обладает стереоспецифичностью, поэтому в результате этой реакции образуется еноил-ацилКоА. В процессе окисления ФАД восстанавливается и передает электроны в дыхательную цепь. Окисление, катализируемое ацил-КоА дегидрогеназой аналогично окислению сукцината в ЦТК. 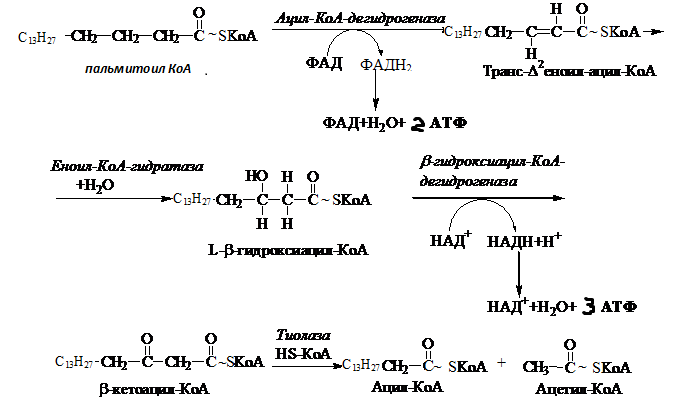 2. Гидратация. Еноил-КоА-гидратаза присоединяет молекулу воды и образуется β-гидроксиацил-КоА. 3. Дегидрирование. β-гидроксиацил-КоА-дегидрогеназа (НАД-зависимая дегидрогеназа) катализирует второе окисление. Образуется β-кетоацил-КоА и НАДНН+. 4. Под действием тиолазы (ацил-КоА-ацетилтрансфераза) с использованием свободного HS-KoA β-кетоацил-КоА расщепляется до ацетил-КоА и ацил-КоА, укороченного на два углеродных атома. Окисление ацетил-КоА в ЦТК. Ацетил-КоА вступает в цикл трикарбоновых кислот, конденсируясь с оксалоацетатом. При окислении ацетильной группы ацетил-КоА в ЦТК и сопряженных цепях переноса электронов образуется 12 АТФ. Укороченный ацил-КоА вступает в следующий цикл -окисления. Энергетический баланс окисления жирных кислот Расчет выделяемой энергии при окислении жирной кислоты производится по формуле: [5 *(n/2 - 1) + n/2 * 12] - 2 где: 5 - число молекул АТФ, образуемое при одном цикле -окисления; n - число атомов углерода в жирной кислоте; n/2-1 - число циклов -окисления; n/2 - число молекул ацетил-КоА; 12 - число молекул АТФ при полном окислении ацетильной группы молекулы ацетил-КоА; - 2 – количество высокоэнергетических фосфатов, используемых на активацию жирной кислоты (при активации АТФ распадается на АМФ и пирофосфат). Количество АТФ, образующихся при окислении пальмитиновой кислоты с учетом энергии, используемой на активацию: [5*(16/2 -1) + 16/2*12]-2= 129 АТФ По В.Эллиот и Д.Эллиот коэффициент полезного действия -окисления жирных кислот равен 33%. Теоретически полное окисление 256 г пальмитиновой кислоты дает 45 кг АТФ. Разумеется, такого количества АТФ никогда не бывает из-за постоянного расходования АТФ на энергетические нужды организма. Регуляция окисления жирных кислот При регуляции окисления первостепенное значение имеет доступность жирных кислот. Поступление жирных кислот определяется содержанием жиров в пище и скоростью липолиза эндогенных липидов. β-гидроксиацил-КоА дегидрогеназа ингибируется высоким отношением [НАДН]/[НАД+]; тиолаза ингибируется ацетил-КоА. Концентрация малонил-КоА, первого промежуточного продукта биосинтеза жирных кислот из ацетил-КоА, повышается, если поступает избыток углеводов с пищей; глюкоза не может быть окислена или депонирована в виде гликогена и превращается в жирные кислоты для синтеза триацилглицеролов. Малонил-КоА ингибирует карнитин-ацилтрансферазу I и, следовательно, ингибируется окисление жирных кислот. Окисление глицерола При липолизе триацилглицеролов жировых клеток или ХМ и ЛПОНП кроме жирных кислот образуется глицерол (глицерин). Глицерол свободно транспортируется кровью. В почках, печени, лактирующих молочных железах имеется фермент глицеролкиназа, который в цитозоле катализирует фосфорилирование глицерола в глицерол-3-фосфат. Стадии окисления: Окисление до ПВК Окислительное декарбоксидирование ПВК ЦТК +тканевое дыхание   Рис. Окисление глицерола Энергетический баланс окисления глицерола до СО2 и Н2О: 1)- на стадии глицеролкиназы - 1 АТФ; - на стадии -глицерофосфатдегидрогеназы + 3 АТФ;  - на стадии дегидрогеназы ГАФ НАДН+Н+ + 3 АТФ; - на стадии дегидрогеназы ГАФ НАДН+Н+ + 3 АТФ; - на стадии глицераткиназы + 1 АТФ субстратное - на стадии пируваткиназы + 1 АТФ фосфорилирование; Сумма 7АТФ 2) при окислительном декарбоксилировании пирувата в ацетил-КоА НАДН+Н+ + 3 АТФ 3) окисление ацетил-КоА в ЦТК и сопряженных цепях переноса электронов + 12 АТФ. Итого: 7+3+12 = 22 АТФ Биологической значение окисления ВЖК и глицерола Окисление ВЖК дает большой выход энергии При окислении ВЖК и глицерола образуется целый ряд ключевых метаболитов – ПВК, 3-ФГА, Ацетил-КоА, которые могут вовлекаться в обмен углеводов. Ацетил-КоА, образующийся при окислении ВЖК и глицерола, используется для синтеза ХС, кетоновых тел и др. соединений. |