2 коллоквиум по БХ. Катаболтм(дисеималяция)
 Скачать 3.5 Mb. Скачать 3.5 Mb.
|

|
Гликогенозы - болезни накопления гликогена (11 типов) 1 тип - болезнь Гирке: недостаточность глюкозо-6- фосфатазы:Гепато-и реномегалия,лактатный ацидоз, гипогликемия, гиперлипидемия, кетоз. 2 тип - болезнь Помпе: отсутствие лизосомной а-1,4-глюкозидазы (кислой а- глюкозидазы, у-амилазы) Поражение всех органов (генерализованный гликогеноз), задержка роста, мышечная гипотония, макроглоссия, гипертрофия миокарда (шаровидное сердце) Агликогеноз - нарушение синтеза гликогена Недостаточность гликогенсинтазы - Гипогликемия, кетоз, ранняя смерть 5) разобщители окислительного фосфорилирования. Это вещества, которые подавляют окислительное фосфорилирование, не влияя при этом на процесс переноса электронов дыхательной цепью. Механизм действия разобщителей сводится к тому, что, являясь липофильными веществами, они обладают способностью связывать протоны и переносить их в матрикс, минуя протонный канал Н+ АТФ -синтазы. Выделяющаяся при переносе электронов энергия рассеивается в виде тепла. Различают разобщители: -естественные (продукты перекисного окисления липидов, жирные кислоты с длинной цепью, белки термогенины бурой жировой ткани, большие дозы йодсодержащих гормонов щитовидной железы); -искусственные (динитрофенол, производные витамина К, некоторые антибиотики). 6) все верно,окситиамин-антагонист тиамина.он предотвращает проявление действия тиамина Билет 15 1) Ферменты, которые катализируют эти реакции, — дегидрогеназы (ДГ). Это двухкомпонентные ферменты; они делятся на пиридиновые (ПДГ), которые в качестве кофермента используют производные витамина РР — НАД и НАДФ+, и флавиновые, которые в качестве кофермента используют производные витамина В2 — ФМН и ФАД.  НАД+: Никотинамид-рибоза-фосфат Аденин-рибоза-фосфат НАДФ+: Никотинамид-рибоза-фосфат  Аденин-рибоза-фосфат фосфат ФАД: Изоаллоксазин-рибитол-фосфат Аденин-рибоза-фосфат ФМН: Изоаллоксазин-рибитол-фосфат Значение: в ходе дегидрогеназных реакций субстраты окисляются, а коферменты восстанавиваются. Восстановленные коферментные формы НАДН*Н+ и ФАДН2 поставляют водород в ДЦ, где происходит его ок-е до воды и синтез АТФ. 2) ) Катаболическая функция цикла Кребса: конечный путь окисления большинства органических веществ; основной источник запасания энергии в клетке в форме АТФ. Т.к. а-кетоглутарат состоит из 5 атомов С,то до образования ЩУК происходит одно декарбоксилирование образуется два НАДН*Н+ ; один ФАДН2; один АТФ два НАДН*Н+=> 2*2,5-5 моль АТФ; одинФАДН+=> 1*1,5=1,5 моль АТФ; В общем, 1,5+5+1=7,5 моль АТФ и остаётся 4 атома С, за один цикл Кребса происходит два декарбоксилирования, данное вещество проходит дополнительно два полных цикла Кребса. Т.к за один цикл выделяется 10 моль АТФ, а у нас 2 оборота цикла, значит выделяется 20 моль АТФ и в сумме получается 7,5+20-27,5 моль АТФ. 3) 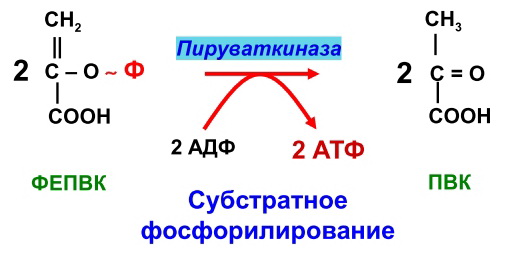 Ключевые ферменты: >Гексокиназа (глюкокиназа) >ФФК-1 >Пируваткиназа Виды регуляции: ^Индукция синтеза ключевых ферментов ^Аллостерическая регуляция ^Ковалентная модификация (фосфорилирование/дефосфорилирование) 4) Распад гликогена в мышцах происходит при мышечных сокращениях, а в печени — при голодании и в перерывах между приёмами пищи. Основной механизм гликогенолиза — фосфоролиз (расщепление а-1,4-гликозидных связей с участием фосфорной кислоты и гли- когенфосфорилазы) 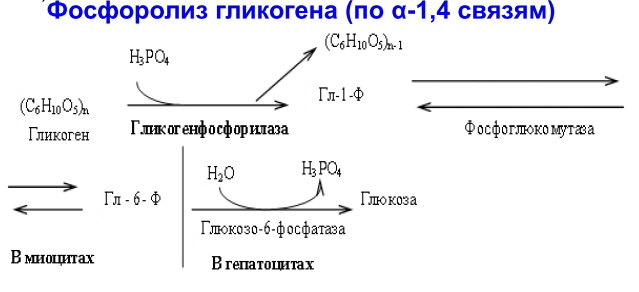 Глюкагон и адреналин стимулируют гликогенолиз, инсулин — тормозит. Регуляция гликогенолиза осуществляется на уровне гликогенфосфо- рилазы. Глюкагон и адреналин активируют (переводят в фосфорилированную форму) глико- генфосфорилазу. Глюкагон (в гепатоцитах и кардиомиоцитах) и адреналин (в миоцитах) активируют гликогенфосфорилазу по каскадному механизму через посредника — цАМФ. Связываясь со своими рецепторами на цитоплазматической мембран клеток, гормоны активируют мембранный фермент аденилатциклазу. Аденилатциклаза нарабатывает цАМФ, который активирует протеинкиназу А, и запускается каскад превращений ферментов, заканчивающийся активацией гликогенфосфорилазы. Инсулин инактивирует, то есть переводит в не- фосфорилированную форму, гликогенфосфорилазу. Мышечная гликогенфосфорилаза активируется АМФ по аллостерическому механизму. Таким образом, гликогенез и гликогенолиз координированно регулируются глюкагоном, адреналином и инсулином. 5)В процессе аэробного ок-я глюкозы образ НАД\ФАД-зависимые субстраты, которые передают протоны и электроны на комплексы ДЦ. Электроны транспортируются по этим комплексам к кислороду, а протоны выходят в межмембр пространство в 1,3,4 комплексах ДЦ. Эти пункты – пункты сопряжения или фосфорилирования. Они создают электрохим потенц, энергия которого используется 4-ым комплексом ВММ для синтеза АТФ 6) -,+,- с потреблением органических субстратов в р-ях дегидрирования субстраты окисл-ся,а коферменты восст-ся. Основным поставщиком субстратов явл-ся центральные метаболические пути,локализованные в митохондриях.Восстановленные коферменты передают водород в дахательной цепи ВММ,где происходит его окисл-ие до воды и синтез АТФ. Билет 16 оксидазный путь - Большая часть кислорода, потребляемого клеткой (около 80 %), используется, как указано, в митохондриях с участием цитохромоксидазы. При этом происходит: полное восстановление кислорода субстрат не реагирует с кислородом непосредственно путь дает клетке энергию в виде АТФ Помимо цитохромоксидазы существуют другие оксидазы (ФМН и ФАД-зависимые), которые катализируют реакции окисления веществ с образованием перекиси водорода. В микросомах происходит другой путь- оксигеназный путь окисления . Он не дает клетке энергии, кислород включается в субстрат с образованием новой гидроксильной или карбоксильной группы. Этот путь происходит в основном в мембранах эндоплазматического ретикулума (микросомах). Путем микросомного окисления осуществляется альфа- и w-окисление жирных кислот, синтез ненасыщенных жирных кислот, стероидов. Таким путем обезвреживаются ксенобиотики, т. е. чужеродные для организма вещества (лекарства, ядохимикаты, косметические препараты). Ферменты, осуществляющие такое окисление, называются оксигеназами. Монооксигеназные системы представляют собой короткие цепи переноса электронов и протонов, источником которых служит чаще всего восстановленный НАДФ+, реже НАД+ или аскорбиновая кислота. Активатором кислорода при этом является цитохром Р450 — одноцепочечный хро- мопротеин с молекулярной массой 50 кДа. 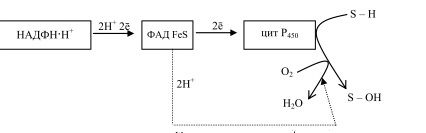 Смысл такого процесса заключается в том, что ксенобиотики, которые обычно гидрофобны, гидроксилируясь, становятся более гидрофильными, что способствует их обезвреживанию и выведению из организма с желчью или мочой. С участием микросомных систем осуществляется также биосинтез стероидов, желчных кислот, витамина Д3. + свободно-радикальный путь 2) Катаболическая функция цикла Кребса: конечный путь окисления большинства органических веществ; основной источник запасания энергии в клетке в форме АТФ. Образуется три НАДН*Н+; один ФАДН2; и один АТФ в ходе субстратного формирования. При окислении НАД- зависимых коэффициент фосфорилирования (Р\О)=2,5 => три НАДН*Н+ дадут 7,5 моль АТФ. Р/0 при окислении ФАД Н - зависимых субстратах в 1,5 => 1 моль ФАДН2 даст 1,5 моль АТФ. В сумме 1+1,5+7,5=10 моль АТФ 2 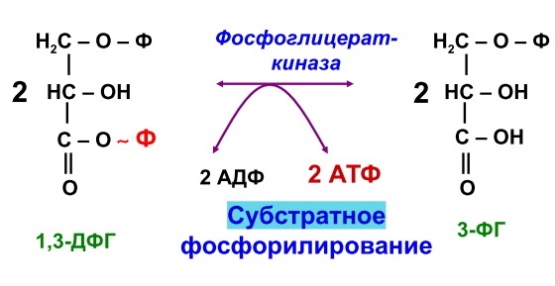 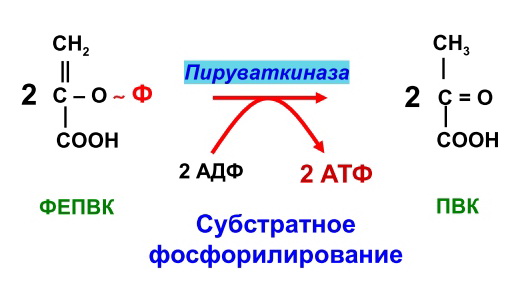 )Энергетический баланс гликолиза — две молекулы АТФ на одну молекулу глюкозы. На 1 этапе гликолиза расходуются две молекулы АТФ для активирования субстрата (в гексокиназной и фосфофруктокиназной реакциях). На 2 этапе образуются четыре молекулы АТФ (в фосфоглицераткиназной и пируваткиназной реакциях). Синтез АТФ осуществляется путем субстратного фосфорилирования. Анаэробный гликолиз основной источник энергии в эритроцитах, злокачественных клетках, интенсивно работающей мышце (кратковременно) В переваривании углеводов участвуют спец ферменты: а-Амилаза слюны,Панкреатическая а-амилаза,дисахаридазы,Мальтаза,Лактаза ,Изомальтаза/Сахараза,Трехалаза,а-Декстриназа (терминальная декстриназа) Всасывание глюкозы (галактозы) происходит в два этапа I этап — транспорт глюкозы из полости тонкого кишечника в энтероциты. Осуществляется по двум механизмам: натрий-независимый транспорт с участием ГЛЮТ 5 (транспортер глюкозы 5); натрий-зависимый транспорт с участием Na -глюкозного транспортёра. II этап — транспорт глюкозы из энтероцитов в капилляры портальной венозной системы (натрий-независимый транспорт с участием ГЛЮТ 2). 5)НАД-завис субстраты = 7,5 АТФ ФАД-завис субстраты = 1,5АТФ 6) +,-,-. Распад галактозы зависит от галл-1-Ф-уридилтрансферазы(гал-1-Ф до гл-1-Ф) Билет 17 1) Субстратное фосфорилирование - синтез АТФ за счёт энергии гидролиза макроэргической связи субстрата (пример: фосфоглицераткиназная и пируватк иназная реакции анаэробного гликолиза). В цитозоле и мотохондриях. 2)  АТФ свободного гликолиза = 30-32 АТФ глюк из мыщц = 31-33 3) Возвращение лактата в метаболический фонд углеводов. Лактат, образующийся в процессе анаэробного окисления глюкозы в эритроцитах и скелетных мышцах, транспортируется кровью в печень и превращается в гепатоцитах в глюкозу. Это так называемый межорганный цикл Кори . Предотвращение лактатного ацидоза, то есть в ходе глюконеогенеза лактат крови превращается в глюкозу.  Энергетические затраты на синтез глюкозы и лактата =4АТФ + 2 ГТФ(6АТФ) 4) Пищевые волокна. Некрахмальные полисахариды состоят из гетерогенной группы углеводных соединений (клетчатка, пектины, гемицеллюлоза, камеди). Основной полисахарид пищевых волокон — клетчатка (целлюлоза). Некрахмальные полисахариды не усваиваются организмом человека, так как в пищеварительном тракте отсутствуют ферменты их гидролиза. Роль пищевых волокон в питании человека: 1. Поддерживают эубактериоз кишечника. Стимулируют перистальтику кишечника. Связывают воду и удерживают её в кишечнике. Создают давление на стенки кишечника и желудка. Являются энтеросорбентами. Образуют гели, что оказывает защитное действие на слизистую кишечника. Создают более длительное чувство насыщения. Лактулоза – восстанавл дисахарид, состоящий из гал и фр связанных бета-1,4-гликозидной связью Целлюлоза(клетчатка) – линейный гомополисахарид, состоящий из бетаДглюкопираноз связанных бета-1,4-гликозидной связью Пектин свзывает тяжелые Ме, радионуклиды и не дает им всасываться. 5) Цитохромы, входящие в состав дыхательной цепи, представляют собой железосодержащие белки, простетическая группа которых представлена гемом. Цитохромы могут переносить только электроны за счет атома железа с переменной валентностью, входящего в состав гема: атомы железа и меди в составе цитохромов; цитохромы–это хромопротеины. Классы: цит. А, цит. В, цит. С (различаются по строению гема). Цит. А и А3, цит. С и С1.. Переносят только электроны: Fe3+ + ē ↔ Fe2+ Cu2+ + ē ↔ Cu+ Включаются в ДЦ в опред последовательности В → С1 → С → А → А3 Цитохром с-оксидаза. Переносит электроны с цитохрома с непосредственно на кислород. Цитохромы а и а3, помимо атомов железа, содержат атомы меди, поэтому этот комплекс одновременно осуществляет полное (4-электронное) восстановление молекулы кислорода. Энергия переноса электронов используется на перекачивание в межмембранное пространство протонов. 6) +,-,-. ПВК (уменьшается) ,лактат (увеличивается) ,т.к. в процессе ->гликолиза ПВК лактат Билет 18 1)В микросомах происходит путь оксигеназный окисления . Он не дает клетке энергии, кислород включается в субстрат с образованием новой гидроксильной или карбоксильной группы. Этот путь происходит в основном в мембранах эндоплазматического ретикулума (микросомах). Путем микросомного окисления осуществляется альфа- и w-окисление жирных кислот, синтез ненасыщенных жирных кислот, стероидов. Таким путем обезвреживаются ксенобиотики, т. е. чужеродные для организма вещества (лекарства, ядохимикаты, косметические препараты). Ферменты, осуществляющие такое окисление, называются оксигеназами. Монооксигеназные системы представляют собой короткие цепи переноса электронов и протонов, источником которых служит чаще всего восстановленный НАДФ+, реже НАД+ или аскорбиновая кислота. Активатором кислорода при этом является цитохром Р450 — одноцепочечный хро- мопротеин с молекулярной массой 50 кДа. Смысл такого процесса заключается в том, что ксенобиотики, которые обычно гидрофобны, гидроксилируясь, становятся более гидрофильными, что способствует их обезвреживанию и выведению из организма с желчью или мочой. С участием микросомных систем осуществляется также биосинтез стероидов, желчных кислот, витамина Д3. + свободно-радикальный путь 2) 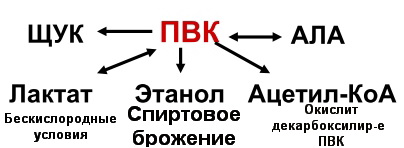 При энергодефиците-окислит декарбоксилир-е ПВ К 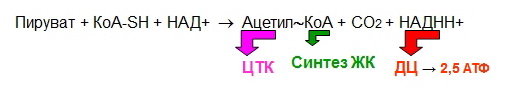 3)Регуляция 1. Пируваткарбоксилаза активируется АМФ и ацетил-КоА Глюкагон -> цАМФ > Протенкиназа А—> фосфорилирование фосфофруктоки-назы 2-> уменьшение образования фруктозо-2,6 диф-та-> стимуляция глюконеогенеза  Кортизол оказывает катаболическое действие на мышечную и лимфоидную системы (распад белков) и анаболическое - на печень (индукция синтеза ферментов глюконеогенеза), канальцы почек, слизистую кишечника 4) Существует группа белков-переносчиков глюкозы (ГЛЮТ), сходных по структуре, но различающихся по участию в транспорте глюкозы (изоформы собственных транспортеров глюкозы). Они локализованы в плазматических мембранах всех клеток и участвуют в транспорте глюкозы (ускоряют транспорт) по градиенту её концентрации. 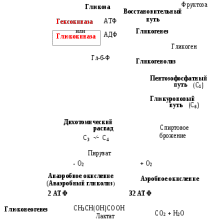 В плазматических мембранах всех клеток есть ГЛЮТ ≈ 500 АК В плазматических мембранах всех клеток есть ГЛЮТ ≈ 500 АКГЛЮТ 1 (эритроциты, плацента, почки, толстый кишечник) ГЛЮТ 2 (печень, поджелудочная железа (Р-клетки), почки, тонкий кишечник) ГЛЮТ 3 (мозг, плацента, почки, скелетные мышцы плода) ГЛЮТ 5 (тонкий кишечник) ГЛЮТ4 (жировая ткань, скелетные мышцы, сердечная мышца) Инсулин стимулирует поступление глюкозы в адипоциты, миоциты и кардиомиоциты, увеличивая количество ГЛЮТ 4 в плазматических мембранах этих клеток. Инсулиннезависимый транспорт глюкозы (печень, мозг, форменные элементы крови, мозговой слой почек, зародышевый эпителий семенников) 5) I. НАДН-убихинон-оксидоредуктаза. Принимает электроны и протоны от НАДН-Н+; протоны выбрасываются в межмембранное пространство, электроны передаются на КоQ. ФМН: Изоаллоксазин-рибитол-фосфат В состав комплекса также входят железосодержащие белки (FeS-белки), котор разделяют потоки протонов и электронов.Элетроны от ФМН2 перенос к внутр пов ВММ, а протоны к внеш пов ВММи далее в ММП(выбрас 4Н+) 6) )+,-,-. Кортизол стимулирует глюконеогенез,а повышает активность печени гликонесинтазы(т.е.переводит её в активную нефосфорилированную форму). Инсулин инактивирует глюкагон |