Клеточная теория. Клеточная теория. Химический состав клетки
 Скачать 139.5 Kb. Скачать 139.5 Kb.
|

|
Аденозинтрифосфат (АТФ). Входит в состав любой клетки, где он выполняет одну из важнейших функций — на копителя энергии. Молекулы АТФ состоят из азотистого основания аденина, углевода рибозы и трех молекул фосфорной кислоты. Не устойчивые химические связи, которыми соединены молекулы фос форной кислоты в АТФ, очень богаты энергией (макроэргические связи): при разрыве этих связей энергия высвобождается и исполь зуется в живой клетке для обеспечения процессов жизнедеятель ности и синтеза органических веществ. 3. КЛЕТОЧНЫЕ СТРУКТУРЫ И ИХ ФУНКЦИИ Цитоплазма. Клетка представляет собой целостную живую систему, состоящую из неразрывно связанных между собой цитоп лазмы и ядра (рис. 2). От внешней среды цитоплазма отграничена наружной клеточной мембраной (от лат. membrana — кожица, пленка), называемой плазмалеммой. Она представляет собой тон чайшую (толщина 7—10 нм), но довольно плотную пленку, состоящую почти исключительно из упорядочение расположенных молекул белков и липидов. Наружный и внутренний слои элемен тарной мембраны образованы белковыми молекулами, а между ними находятся два слоя липидов. Молекулы большинства липидов состоят из полярной головки и двух неполярных хвостов. Головки гидрофильны (проявляют сродство к воде), а хвосты гидрофобны (с водой не смешиваются). 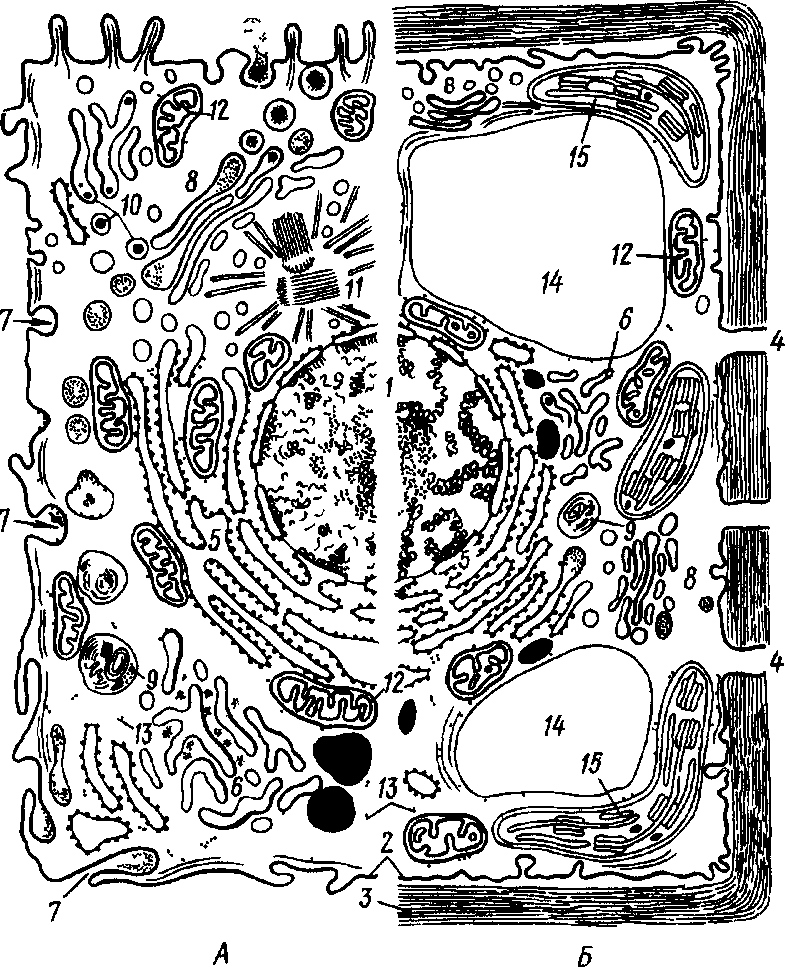 Рис 2 Схема строения эукариотической клетки А — клетка животного, Б — растительная клетка ./ — ядро с хроматином и ядрышками, 2— цитоплазматическая мембрана, 3— клеточная стенка, 4— поры в кле точной стенке, через которые сообщается цитоплазма соседних клеток, 5— шероховатая эндоплазматическая сеть, б— глад кая эндоплазматическая сеть, 7— пиноцитозная вакуоль, 8— аппарат Гольджи, 9— лизосомы, 10— жировые включения, 11— клеточный центр, 12— митохондрия, 13— рибосомы и полирибосомы, 14— вакуоль, 15— хлоропласт Рис. 3. Схема строения цитоплазматической мембраны В двойном липидном слое (I) молекулы липидов направлены своими гидрофильными кон цами (2) к слоям белков (3), состоящих из отдель ных субъединиц Некоторые субъединицы (4) пересекают мембрану) (туннельные белки) Большинство молекул белков располагается на поверхности липидного каркаса с обеих его сторон, а некоторые молекулы проходят через него насквозь, образуя в мембране гидрофильные поры, через которые поступают водорастворимые вещества (рис. 3). Жирорастворимые вещества перемещаются между молекулами белка через липидный слой. Одно из основных свойств клеточных мембран — их избирательная проницаемость (полупроницаемость): одни вещества проходят через них легко, и даже против градиента концентрации, другие с трудом. Благодаря этому свойству наружная клеточная мембрана регулирует проникновение веществ в клетку и выход их из клетки. Внутренняя мембрана клетки — тонопласт — окружает вакуоли. Она устроена так же, как и наружная мембрана. 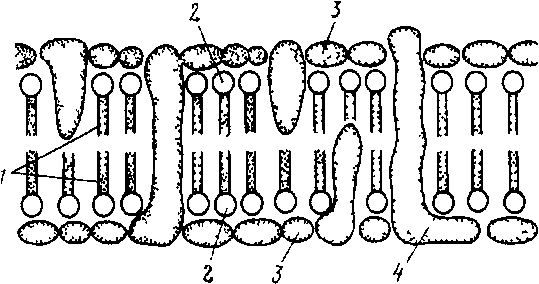 Цитоплазма содержит множество специализированных компонентов, называе мых органеллами. Каж дый вид органелл выполняет определенную функцию. Их согласован ная деятельность обес печивает биохимические реакции, необходимые для жизнедеятельности организма. К органеллам относятся эндоплазматическая сеть, рибосомы, митохондрии, пластиды, лизосомы, комплекс Гольджи, клеточный центр и др. Органеллы взвешены в жидкой среде цитоплазмы, которую назы вают цитоплазматическим матриксом или гиалоплазмой. Это наименее плотная часть клетки, представляющая собой водную кол лоидную систему. Коллоидность определяет способность ее загустевать и превращаться в твердый студень (гель), который, в свою очередь, при определенных условиях разжижается и вновь прев ращается в жидкость (золь). Такой процесс происходит, например, при прорастании семян. Функциональное значение гиалоплазмы как среды, в которую погружены клеточные органеллы, следующее: 1) благодаря вязкости и способности к перемещению гиалоплазма служит основной магистралью для передвижения метаболитов клетки; 2) примыкая к наружной клеточной мембране, она обеспечивает обмен веще ствами между клетками; 3) вступая в непосредственные контакты с мембранами органелл, обусловливает физико-химические и фер ментные связи между ними. Эндоплазматическая сеть. Это разветвленная система каналов и цистерн, ограниченных мембранами, пронизы вающих гиалоплазму. Мембраны, образующие стенки эндоплазматической сети, по структуре сходны с наружной клеточной мембраной. Их толщина в среднем 8 нм, а диаметр каналов 20—50 нм. Каналы заполнены бесструктурной жидкостью — матриксом. Существует два типа эндоплазматической сети — гладкая (агранулярная) и шероховатая (гранулярная). На мембранах первого типа находятся ферменты жирового и углеводного обмена, т.е. на них происходит синтез липидов и углеводов. На мембранах второго типа располагаются рибосомы, придающие мембранам шероховатый вид. Белки, синтезируемые в них, накапливаются в каналах и полостях эндоплазматической сети и затем по ним доставляются к различным органеллам клетки, где используются или сосредоточиваются в цитоплазме в качестве клеточных включений. Кроме того, по каналам эндоплазматической сети вещества транспортируются из клетки в клетку. Рибосомы. Небольшие органеллы диаметром около 22 нм. Они содержат примерно равные количества белка и рибосомальной РНК и лишены мембранной структуры. Каждая рибосома состоит из двух субъединиц различной величины, соединенных между собой. Субъединицы образуются в ядрышках; сборка рибосом осуществляется в цитоплазме. Рибосомы — постоянная составная часть клетки. Часть их располагается в гиалоплазме свободно, другая часть прикреплена к поверхности мембран эндоплазматической сети. Последние функционально более активны. Рибосомы могут располагаться на мембране поодиночке или объединяться в группы по 4—40 единиц, образуя цепочки — полисомы или полирибосомы, в которых отдельные рибосомы связаны между собой нитевидной молекулой мРНК. Рибосомы несколько меньшего размера содержатся в митохондриях и пластидах. Основная функция рибосом — «сборка» белковых молекул из аминокислот. Митохондрии (от греч. митос — нить, хондрион — зерно,гранула). Содержатся во всех аэробных эукариотических клет-ках. Митохондрии впервые обнаружил в мышечных клетках Лёлликер (1850). Митохондрии—микроскопические структуры размером от 0,5(0,3) до 1 мкм в поперечнике и от 2 до 5—7 мкм в длину. Форма их разнообразна: от сферических глыбок (зерны-шек) до телец нитевидной формы. Число митохондрии в клетке ко леблется в широких пределах и зависит от типа тканей и возраста слагающих их клеток. Митохондрии способны перемещаться в клет-ке. При этом они концентрируются преимущественно возле ядра, хлоропластов и других органелл, где процессы жизнедеятельности наиболее интенсивны. Каждая митохондрия окружена двумя мембранами — наружной и внутренней, между которыми находится бесструктурная жидкость матрикс. В клетках животных внутренняя мембрана митохондрии образует на продольной оси многочисленные складки — гребни, а в клетках растений — извилистые трубочки (кристы), увеяичивающие рабочую площадь, на которой расположены окислительные ферменты, участвующие в синтезе АТФ. Промежутки между кристами, или трубочками, также заполнены матриксом. В матриксе митохондрии обычно встречаются более мелкие, чем в гиалоплазме, рибосомы и молекулы ДНК. В митоховдриях осуществляется процесс дыхания. На их внутренних мембранах окисляются пищевые продукты (углеводы, жиры и др.) и накапливается химическая энергия в макроэргических фосфатных связях АТФ. Следовательно, митохондрии можно назвать энергетическими центрами клетки. Число митохондрии в клетке увеличивается путем деления надвое в результате образования перетяжек перпендикулярно их продольной оси. Лизосомы (от греч. лизис — растворяю, сома — тело). Органеллы величиной 0,5—2 мкм. Обнаруживаются в клетках большинства эукариотических организмов, но особенно много их в тех животных клетках, которые способны к фагоцитозу. Они име ют сферическую форму, окружены мембраной и заполнены густо- зернистым матриксом. Лизосомы содержат ферменты, которые, могут разрушать белки, нуклеиновые кислоты, полисахариды, липиды и другие органические соединения при внутриклеточном пищеварении. Число ферментов в лизосомах так велико, что при освобождении они способны разрушить всю клетку. Образно их на зывают «органеллами самоубийства». Большое значение имеет очищение с помощью лизосом полости клетки после отмирания ее протопласта (например, при формировании сосудов и трахеид у растений). По происхождению лизосомы являются производными эндоплазматической сети или аппарата Гольджи. Аппарат Гольджи. Эта структура названа по имени открывшего ее в 1898 г. итальянского ученого К. Гольджи и до недавнего времени признавалась как компонент только животной клетки. Однако с помощью электронного микроскопа выяснилось присутствие аппарата Гольджи почти во всех типах и растительных клеток. Аппарат Гольджи состоит из диктиосом. Каждая диктиосома представляет собой стопку плоских мешочков — цистерн диаметром около 1 мкм и толщиной 20—25 нм, отграниченных мембраной и расположенных параллельно друг другу. Число цистерн в диктиосоме 5—7. От краев цистерн отчленяются пузырьки различного диаметра — пузырьки Гольджи. Вся структура носит полярный ха рактер. Имеются формирующий полюс диктиосомы, где возникают новые цистерны, и секретирующий полюс, где цистерна распадается и образуются секретирующие пузырьки. Такое обновление происходит постоянно и сопровождается повышением активности цистерн по мере продвижения к секреторному полюсу, что выра жается в увеличении числа отчленяемых пузырьков. Новые цистер ны образуются на формирующем полюсе из элементов эндоплазматической сети. В животных клетках аппарату Гольджи уже давно приписыва лась секреторная функция. К настоящему времени для раститель ных клеток выяснено, что диктиосомы являются центром синтеза, накопления и секреции полисахаридов клеточной стенки. Об этом свидетельствует усиление их активности в области «клеточной пластинки», т.е. в той области, где после деления ядра (митоза или мейоза) между двумя только что образовавшимися дочерними ядрами закладывается новая клеточная стенка. При этом пузырьки Гольджи направляются к нужному месту на клеточной пластинке при помощи микротрубочек и здесь сливаются. Их мембраны ста новятся частью новых плазмалемм дочерних клеток, а их содержимое используется для построения стенок этих клеток. В каждой растительной клетке обнаруживается несколько диктиосом. В животных клетках чаще встречается одна диктиосома. Микротрубочки. Это полые цилиндрические органеллы, содержащиеся почти во всех эукариотических клетках. Они представляют собой очень тонкие трубочки (диаметр 24 нм, длина не-сколько микрометров), лишенные мембранной структуры. Их стенки толщиной около 5 нм построены из спирально упакованных глобулярных субъединиц белка — тубулина. Растут микротрубочки с одного конца путем добавления тубулиновых субъединиц. Микротрубочки принимают участие в различных внутриклеточных процессах: входя в состав центриолей и веретена деления ядра, регулируют расхождение хроматид или хромосом, обеспечивая их движение за счет скольжения микротрубочек, входящих в состав базальных телец ресничек и жгутиков, а также, как отмечалось выше, перемещают в нужном направлении пузырьки Гольджи. Клеточный центр. Это органелла, находящаяся вблизи ядра в клетках животных. Она состоит из двух маленьких телец цилиндрической формы (центриолей), расположенных под прямым углом друг к другу. Центриоли содержат ДНК и относятся к самовоспроизводящимся органеллам цитоплазмы. Стенка центриоли состоит из микротрубочек. Центриоли играют важную роль при делении клетки: от них начинается рост микротрубочек, формирующих веретено деления. Органеллы движения клеток. К ним относятся реснички и жгутики (миниатюрные выросты клеток в виде волосков), с помощью которых одноклеточные организмы легко движутся в жидкой среде. Они широко распространены как у одноклеточных, так и у многоклеточных животных. Среди простейших с помощью жгутиков перемещаются жгутиконосцы, а с помощью ресничек — инфузория туфелька. Целый ряд одноклеточных организмов л отдельных клеток животных движется с помощью выпячивания участков тела — ложноножек (амеба, слизевики, лейкоциты). Такой способ передвижения называется амебоидным. У многоклеточных животных и растений с помощью жгутиков передвигаются сперматозоиды и зооспоры. Ядро. Это органелла, где хранится и воспроизводится наследственная информация, определяющая признаки данной клетки и всего организма в целом. Ядро является также центром управления обмена веществ клетки, контролирующим деятельность всех других органелл. Поэтому, если ядро удалить, клетка, как правило, погибает. Форма ядра чаще всего шаровидная или эллипсоидальная, реже линзообразная или веретеновидная. Размер ядра очень изменчив и зависит от вида организма, а также от возраста и состояния клетки. Иногда «рабочая поверхность» ядра увеличивается путем образования лопастей или отростков. В физико-химическом отно шении ядро клетки представляет собой комплекс гидрофильных коллоидов более вязкой консистенции, чем коллоидная система цитоплазмы. По химическому составу ядро резко отличается от остальных органелл высоким (15—30 %) содержанием ДНК и РНК (12,1 %). Почти вся ДНК клетки (99 %) находится в ядре, где она образует комплексы с белками — дезоксирибонуклеопротеиды (ДНП). Основное вещество ядра — сложные белки (протеины). В ядре есть также липиды, вода, ионы Са + и Mg +. Общий план строения ядра одинаков как у растительных, так и у животных клеток. Структура же компонентов ядра существенно изменяется на разных фазах жизненного цикла клетки, что связано с различием выполняемых ядром функций. В связи с этим выделяют три состояния ядра: 1) делящееся ядро, выполняющее функцию передачи наследственной информации от клетки к клетке; 2) ядро, синтезирующее (редупликация) наследственный материал — ДНК (это состояние характерно для ядер в промежутках между де лениями); 3) рабочее ядро живых неделящихся клеток, выполня ющее функцию управления жизнедеятельностью клетки. В ядре различают: 1) ядерную оболочку; 2) хроматин (хромо сомы); 3) одно-два, иногда несколько ядрышек; 4) ядерный сок. Ядерная оболочка. Она имеет малую толщину и поэто му не видна в световом микроскопе. Она состоит из двух мембран, разделенных бесструктурным матриксом, сходным с матриксом ка налов эндоплазматической.сети. Наружная мембрана ядерной обо лочки непосредственно связана с каналами эндоплазматической сети. Поверхность ее покрыта рибосомами. Ядерная оболочка содержит своеобразные структуры — ядерные поры. В них наружная и внутренняя ядерные мембраны слиты по краям. Диаметр пор от 30 до 100 нм. Число пор также колеблется, занимая в зависимости от метаболической активности ядра и вида организма от 10 до 50 % общей площади его поверхности. Поры — это не простые отверстия, а сложные структурные образования, обеспечивающие избирательную проницаемость. Ядерная оболочка контролирует обмен веществ между ядром и цитоплазмой. Из ядер ного сока в гиалоплазму проходят макромолекулы, в том числе предшественники рибосом, и осуществляется транспорт белков в обратном направлении. Хроматин, или хромосомы (от греч. хрома — цвет, сома — тело). Это основной морфологический компонент ядра. Под световым микроскопом на фиксированных и окрашенных препара тах он выявляется в виде сети из тонких длинных нитей, а также , мелких гранул. Исследованиями ядра под электронным микроско пом установлено, что основу хроматина составляют тонкие (10 нм) нити (фибриллы), скрученные в спирали. По химическому составу они представляют собой на 90 % дезоксирибонуклеопротеиды и на 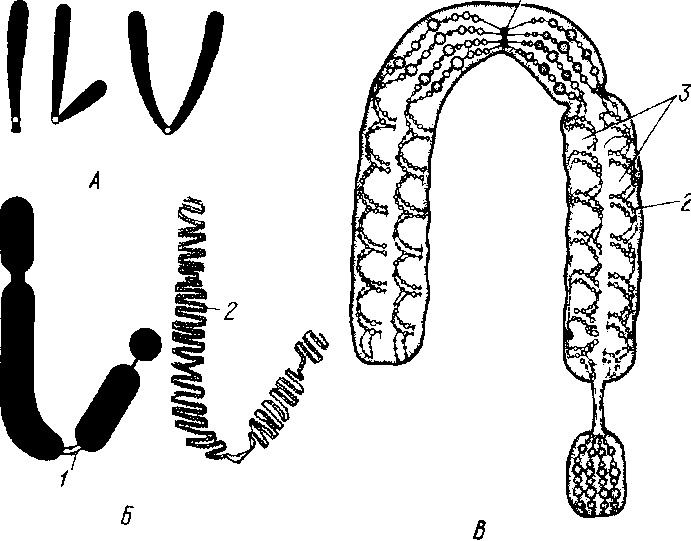 Рис. 5. Строение хромосом. Л— типы хромосом; Б, В— тонкое строение хромосом: /— центромера, 2— спирально закрученная нить ДНК, 3— хроматида 10 % рибонуклеопротеиды. Хроматин — это деспирализованные и гидратированные хромосомы. Таким образом, хромосомы присутствуют в ядре всегда, но в рабочем ядре они обычно не видны, так как находятся в «разрыхленном» состоянии. Хромосомы хорошо видимы в световой микроскоп во время митоза. Для клеток каждого вида характерно постоянное число хромосом определенной величины и формы. Совокупность хромосом называется хромосомным набором. Число хромосом в соматических клетках (от лат. soma — тело) обычно двойное (диплоидное). Оно получается после слияния двух половых клеток, в которых всегда одинарное (гаплоидное) число хромосом. Каждый гаплоидный набор обозначается через п, а диплоидный — через 2n. Размеры и форма хромосом одного гаплоидного набора не одинаковы, но в каждой половой клетке одного вида организма строго повторяется не только число хромосом, но и размеры и форма каждой из них. Естественно, что в диплоидном наборе каждой хромосоме соответствует парная (гомологичная) хромосома, такая же по форме и размерам. Все организмы одного вида имеют одинаковое число хромосом. Так, у мягкой пшеницы их 42, у кукурузы — 20, у коровы — 60, у курицы — 78, а у плодовой мушки дрозофилы -8. Хромосома делящегося ядра имеет вид двойной палочки. Она стоит из двух половин, разделенных узкой щелью вдоль оси хромосомы и называемых хроматидами (рис. 5). Каждая хроматида включает две или несколько спирально закрученных тонких нитей, расположенных параллельно оси хромосомы, называемых хромонемами. Участки наиболее плотных завитков спирали хромонем на зываются хромомерами. Каждая хромосома имеет первичную перетяжку, которая пред ставляет собой неспирализованный участок хромосом, где распо ложена центромера (кинетохор). Перетяжка выглядит как утонченная часть хромосомы. Первичная перетяжка делит хромо сому на две части — два плеча. В зависимости от местоположения перетяжки выделяют три типа хромосом (рис. 5): 1) палочкообразные с одним очень длинным и другим очень коротким, иногда почти незаметным плечом; 2) неравноплечие (плечи неравной длины); 3) равноплечие (плечи равной длины). Иногда хромосома имеет и вторичную перетяжку. Если она рас положена вблизи конца хромосомы и отделенный ею участок не велик, его называют спутником, а несущую его хромосому — спутничной. Расположение и длина перетяжек постоянны для каж дой хромосомы. Вторичная перетяжка — это место, где формируется ядрышко, поэтому ее называют организатором ядрыш ка. Внутреннее строение хромосом, число в ней нитей ДНК ме няются в жизненном цикле клетки. Функции хромосом состоят в синтезе специфических для данного организма нуклеиновых кислот ДНК, хранящих и передающих наследственную информацию в кле точных поколениях, и РНК, управляющих синтезом белков в клет ке. Ядрышки. Размеры и число их более или менее постоянны для одного вида. Форма ядрышка шаровидная, границы неот четливы, так как ядрышки не окружены мембраной и находятся в непосредственном контакте с ядерным соком. Для химического состава ядрышка характеры рибонуклеопротеиды, липопротеиды, фосфопротеиды. Содержание белков в нем весьма велико. Концентрация РНК в ядрышке выше, чем в других частях ядра и в цитоплазме. В нем содержится также немного ДНК. Ядрышки как оформленные тела не существуют постоянно. Они обнаруживаются лишь в неделящемся ядре, а при делении ядра исчезают. В конце деления ядрышки вновь формируются в области вторичных перетяжек некоторых хромосом. Субмикроскопическая структура ядрышек, как правило, универсальна. Они состоят из рыхлого клубочка нитей деспирализованной молекулы ДНК, погруженного в аморфный матрикс. На ДНК ядрышек происходит синтез РНК. Здесь же рРНК объединяется с белком (образование рибонуклеопротеидов), и таким образом формируются и накапливаются предшественники рибосом, Последние через поры в ядерной оболочке переходят в цитоплазму, где и заканчивается их объединение в рибосомы. Таким образом, ядрышки играют важнейшую роль в процессах, предшествующих биосинтезу белков клетки. Ядерный сок (кариоплазма). Представляет собой бесструктурную массу, близкую к гиалоплазме цитоплазмы. Он стоит в основном из простых растворимых белков, а также нуклеопротеидов, гликопротеидов. В нем находится большая часть ферментов ядра. Основная функция ядерного сока — осуществление взаимосвязи ядерных структур (хроматина и ядрышка), но он не является инертной средой, а трансформирует проходящие через него вещества. Таким образом, клетки подавляющего большинства живых организмов имеют сложно устроенное оформленное ядро. Их называют эукариотами. Бактерии и цианеи относят к прокариотам, основной отличительный признак которых — отсутствие ограниченного оболочкой ядра. У них наследственный материал представлен одной-единственной хромосомой, расположенной непосредственно в цитоплазме. Пластиды. Характерны для клеток автотрофных растений. Именно с пластидами связан процесс первичного и вторичного синтеза углеводов. Пластиды различают по окраске: I) бесцветные - лейкопласты; 2) окрашенные в зеленый цвет — хлоропласты; 3) незелёные, окрашенные преимущественно в желто-красные тона,— хромопласты. Все три группы пластид связаны общим происхождением и сходным строением. Предполагают, что пластиды образуются из бесцветных пропластид, которые находятся в клетках зародыша и образовательных тканей. Пропластиды имеют оболоч-ку, состоящую из двух цитоплазматических мембран. Полость заполнена бесструктурным матриксом. Внутренняя мембранная структура пропластид развита очень слабо. Обычно это редкие, часто одиночные цистерны, расположенные без определенной ориентации, иногда это трубочки или пузырьки. Пропластиды могут превращаться в любой из трех видов пластид. |