Коллоквиум. коллок 4 физио 218 К.И.. Коллоквиум 4 Физиология Понятие моторной единицы. Классификация моторных единиц
 Скачать 5.09 Mb. Скачать 5.09 Mb.
|

|
Коллоквиум №4 Физиология 1.Понятие моторной единицы. Классификация моторных единицМоторная единица(МЕ) - является функциональнойединицей скелетноймышцы. МЕ включает всебя группу мышечныхволокон ииннервирующий ихмотонейрон. Классификация Моторные единицы одной мышцы могут быть разными. В зависимости от скорости сокращения моторные единицы разделяют на: • медленные/slow (S-МЕ) • быстрые/fast (F-МЕ) ►быстроутомляемые/fast-fatigable (FF-МЕ) ► и устойчивые к утомлению/fast-fatigue-resistant (FR-МЕ). Характеристика Моторных Единиц Медленные неутомляемые (S-МЕ) имеют низкие морфофункциональные показатели: небольшой мотонейрон, нетолстый аксон, низкий порог раздражения. Содержат много миоглобина, поэтому красные. В них много митохондрий, но мало миофибрилл, большую силу не развивают, хорошо развито кровоснабжение, доставка кислорода, высокая активность окислительных ферментов, снабжаются энергией за счет аэробных процессов. Быстроутомляемые моторные единицы (FF – МЕ) имеют более высокие морфофункциональные показатели: крупный мотонейрон, толстый аксон, скорость проведения 70-120 м/с, более высокий порог раздражения, много миофибрилл, гликогена, мало митохондрий, миоглобина, хуже капиллярное кровоснабжение, преобладают гликолитические ферменты. Аксон разветвляется на большое количество концевых веточек, иннервирующих большую группу мышечных волокон. Быстрые устойчивые к утомлению (FR –МЕ).Они включают сильные, быстро сокращающиеся волокна, обладающие большой аэробной выносливостью. Мышцы, составляющие данную моторную единицу, способны выполнять быстрые сокращения без заметного утомления. 2.Моторный синапс, его строение и особенности передачи возбуждения. Нервно-мышечный синапс — эффекторное нервное окончание на скелетном мышечном волокне. Нейромедиатором в этом синапсе является ацетилхолин. В этом синапсе нервный импульс превращается в механическое движение мышечной ткани. Передача возбуждения в нервно-мышечном синапсе. 1. электрический (нервный импульс) – достижение ПД концевой веточки аксона, деполяризация терминали и выделение ацетилхолина (АХ) в синаптическующель; 2. Химический – диффузия АХ к постсинаптической мембране и образование на ней его комплекса с холинорецептором; 3. Электрический – увеличение ионной проницаемости постсинаптической мембраны, возникновение локального потенциала (потенциала концевой пластинки, ПКП), развитие ПД мышечного волокна. ОСОБЕННОСТИ НЕРВНО-МЫШЕЧНОГО СИНАПСА 1. Нервно-мышечный синапс образован окончанием аксона моторного нейрона и мышечным волокном; 2. Медиатор – ацетилхолин; 3. Постсинаптическая мембрана (часть сарколеммы) имеет множество инвагинаций; 4. На постсинаптической мембране комплекс АХ+холинорецептор формирует ПКП (потенциал концевой пластинки). 3. Структурно-функциональные особенности скелетно-мышечных волокон.Механизм мышечного сокращения и расслабления. 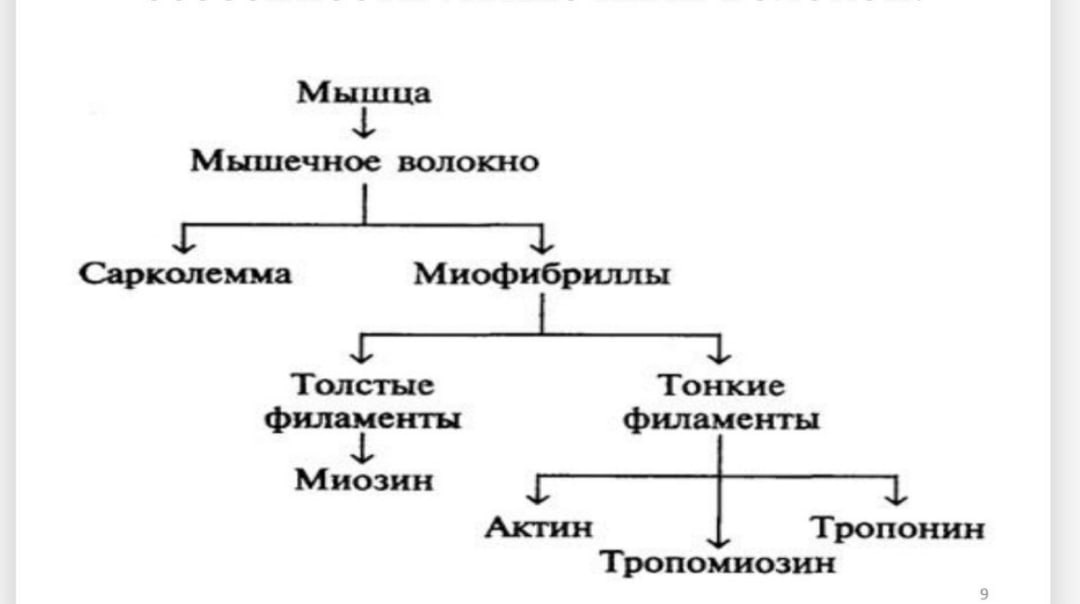 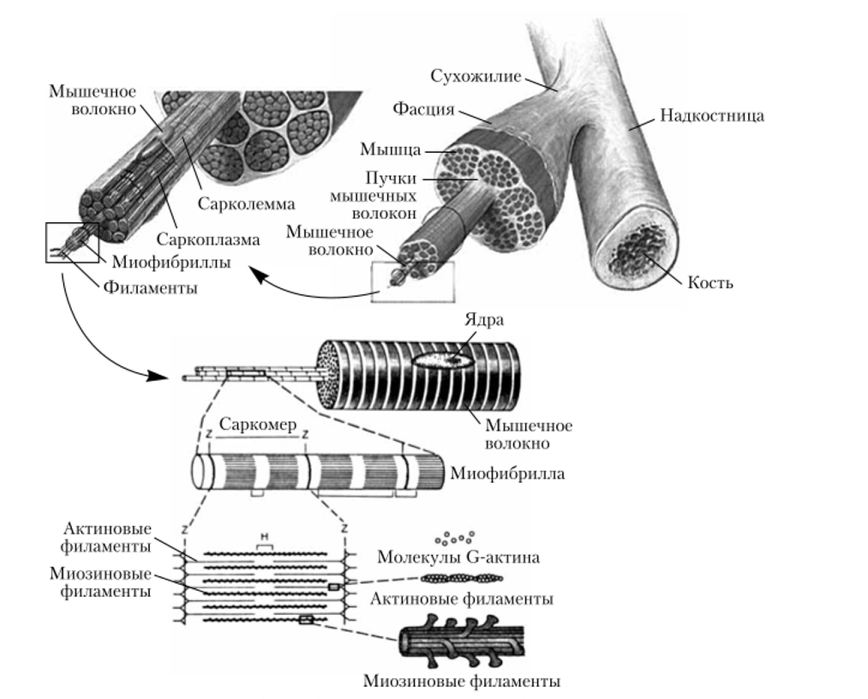      4.Одиночное мышечное сокращение, его фазы. Изменение возбудимости скелетной мышцы в различные фазы одиночного сокращения. При нанесении на двигательный нерв или мышцу одиночного порогового или сверхпорогового раздражения, возникает одиночное сокращение. При его графической регистрации, на полученной кривой можно выделить три последовательных периода: 1) Латентный период: это время от момента нанесения раздражения до начала сокращения. Его длительность около 1-2 мс. Во время латентного периода генерируется и распространяется ПД, происходит высвобождение кальция из СР, взаимодействие актина с миозином и т.д. 2) Период укорочения: в зависимости от типа мышцы (быстрая или медленная) его продолжительность от 10 до 100 мсек., 3) Период расслаблени: его длительность несколько больше, чем укорочения (0,05-0.06 сек.) В режиме одиночного сокращения мышца способна работать длительное время без утомления, но его сила незначительна. Поэтому в организме такие сокращения встречаются редко, например, так могут сокращаться быстрые глазодвигательные мышцы, мышцы сгибателей пальцев. Чаще одиночные сокращения суммируются. Суммация – это сложение двух последовательных сокращений при нанесении на нее двух пороговых или сверхпороговых раздражений, интервал между которыми меньше длительности одиночного сокращения, но больше продолжительности рефракторного периода. Виды: - неполная суммация: возникает в том случае, если повторное раздражение наносится на мышцу, когда она уже начала расслабляться - полная возникает тогда, когда повторное раздражение действует на мышцу до начала периода расслабления, т.е. в конце периода укорочения. Амплитуда сокращения при полной суммации выше, чем неполной. Если интервал между двумя раздражениями еще больше уменьшить, например, нанести второе в середине периода укорочения, то суммации не будет, потому что мышца находится в состоянии рефрактерности. 5.Танатус,его виды и механизм образования.Оптимум и пессимум частот(Н.Е. Введенский). Изменение силы сокращения наблюдают при высокочастотной ритмической стимуляции скелетных мышц. Возникающее в этом случае сильное и длительное сокращение носит название «тетанус». Если скелетную мышцу раздражать такой частотой ритмических импульсов, при которой каждое последующее раздражение будет попадать в период расслабления мышцы от предыдущего раздражения, то возникает так называемая неполная суммация одиночных мышечных сокращений и формируется зубчатый тетанус. При этом сила сокращения постепенно растет, а на кривой сокращения фиксируют характерные зубцы и западения. Если частоту раздражения увеличить до такой степени, что каждое последующее раздражение будет попадать в период укорочения или развития мышечного напряжения от предыдущего раздражения, то происходит так называемая полная суммация одиночных сокращений. В этом случае возникает гладкий тетанус, при котором сила сокращения возрастает быстрее, плавно и до более значительных величин. Механизм возникновения тетанических сокращений. При нанесении второго стимула в период укорочения или развития мышечного напряжения происходит суммация двух следующих друг за другом сокращений и результирующий ответ по амплитуде становится значительно выше, чем при одиночном стимуле; если мышечное волокно или мышцу стимулировать с такой частотой, что повторные стимулы будут приходиться на период укорочения, или развития напряжения, то происходит полная суммация единичных сокращений и развивается гладкий тетанус. Тетанус — сильное и длительное сокращение мышцы. Полагают, что в основе этого явления лежит повышение концентрации кальция внутри клетки, что позволяет осуществляться реакции взаимодействия актина и миозина и генерации мышечной силы поперечными мостиками достаточно длительное время. При уменьшении частоты стимуляции возможен вариант, когда повторный стимул наносят в период расслабления. В этом случае также возникнет суммация мышечных сокращений, однако будет наблюдаться характерное западение на кривой мышечного сокращения — неполная суммация, или зубчатый тетанус. При тетанусе происходит суммация мышечных сокращений, в то время как ПД мышечных волокон не суммируются Оптимум (наилучший) – такая сила и частота раздражителя, которая обусловливает максимальную амплитуду мышечного сокращения (гладкий тетанус). Механизм: концентрация кальция увеличивается в пре- и постсинаптической мембранах, это увеличивает секрецию медиатора, повышение ПКП (ВПСП). Пессимум (наихудший) – такая чрезмерная сила или частота раздражителя, которая вызывает резкое уменьшение амплитуды мышечного сокращения или даже отсутствие реакции мышцы. Механизм: накопление медиатора в щели ведет к уменьшению секреции медиатора из пресинаптической мембраны и к стойкой деполяризации постсинаптической мембраны, к снижению чувствительности и родства к рецепторам постсинаптической мембраны медиатора. 6.Сила и работа мышц, факторы ее определяющие. Сила скелетной мышцы зависит от многих факторов: 1. От числа ДЕ, возбуждаемых в данный момент времени. 2. От синхронности работы ДЕ 3. От частоты ПД «бегущих» по данным аксонам к соответствующим мышечным волокнам. 4. От исходной длины. Работа скелетной мышцы совершается за счет согласованного изменения тонуса (напряжения) и длины мышцы во время сокращения. Виды работы скелетной мышцы: • динамическая преодолевающая работа совершается, когда мышца, сокращаясь, перемещает тело или его части в пространстве; • статическая (удерживающая) работа выполняется, если благодаря сокращению мышцы части тела сохраняются в определенном положении; • динамическая уступающая работа совершается, если мышца функционирует, но при этом растягивается, так как совершаемого ею усилия недостаточно, чтобы переместить или удержатьчасти тела Во время выполнения работы мышца может сокращаться: • изотонически – мышца укорачивается при постоянном напряжении (внешней нагрузке); изотоническое сокращение воспроизводится только в эксперименте; • изометрически – напряжение мышцы возрастает, а ее длина не изменяется; мышца сокращается изометрически при совершении статической работы; • ауксотонически– напряжение мышцы изменяется по мере ее укорочения; ауксотоническое сокращение выполняется придинамической преодолевающей работе. 7.Утомление, его признаки.Теории утомления и способы снятия утомления. Утомление – физиологическое состояние мышцы, которое развивается после совершения длительной работы и проявляется снижением амплитуды сокращений, удлинением латентного периода сокращения и фазы расслабления. Причинами утомления являются: истощение запаса АТФ, накопление в мышце продуктов метаболизма, недостаток притока кислорода. При совершении организмом мышечной работы утомление первоначально развивается на уровне синапсов ЦНС и нейро-мышечных синапсов, затем на уровне нейронов экстрапирамидной и пирамидной системы, нейронов коры.  8.Двигательные, эффекторные нейроны спинного мозга – мотонейроны. Находятся в передних рогах спинного мозга. Различают: • альфа-мотонейроны • гамма-мотонейроны Альфа – мотонейроны – крупные клетки спинного мозга, их аксоны образуют нервномышечные синапсы с экстрафузальными мышечными волокнами (МВ) скелетной мышцы и участвуют в формировании нейромоторных единиц. Его медиадором явл ацитилхолин Бывают альфа1 (передают импульсы на экстрофузальныемыщечные волокна, белые и обеспечивают быстрое сокращение,высокая лабильность ) и альфа 2(медленные. Низкая лабильность, иннерв. Красные волокна.) Гамма – мотонейроны – значительно меньше альфа мотонейронов, осуществляют двигательную иннервацию интрафузальныхМВ(мыш волокна) в составе мышечных веретен.(проприорецепторов) высокая лабильность, но низкая скорость проведения возбуждения. Находятся между альфа мотонейронами. поддерживают тонус скелет мыщц, На одном мотонейроне число синапсов может достигать 10-20тыс 9.Проприорецепторы скелетных мышц, их роль в обеспечении координации рефлекторных актов. Проприоцепторы (от лат. proprius —собственный), специализированные чувствительные нервные окончания из группы механорецепторов, расположенные в опорнодвигательном аппарате (скелетные мышцы, сухожилия, связки) и реагирующие на сокращение и напряжение или расслабление и растяжение мышц. Относятся к первичным рецепторам – чувствительные аксоны афферентного нейрона. Они медленно адаптирующиеся. Рефлексы с проприорецепторов участвуют в формировании акта ходьбы и регуляции мыщечного тонуса, что важно при поддержании позы тела. К проприорецепторам относятся: • мышечные веретена, находящиеся среди мышечных волокон, возбуждаются при их удлинении, основные элементы – интрафузальные МВ, нервные волокна и капсула; • тельца Гольджи и мацони, расположенные всухожилиях, возбуждаются при сокращении мышечных волокон; -суставные рецепторы • пачиниевы тельца, находящиеся в фасциях, покрывающих мышцы, в сухожилиях, связках и периосте, возбуждаются при давлении. 10. Роль гамма – эфферентного контроля в поддержании тонуса скелетных мышц. Степень сокращения веретена регулируется так называемой гамма-системой — гаммаэфферентными нервными волокнами. Каждый раз, когда сигналы передаются от двигательной коры или от любой другой области головного мозга к альфа мотонейронам, в большинстве случаев одновременно стимулируются гаммамотонейроны, что называют коактивациейальфа- и гамма-мотонейронов. Это ведет к одновременному сокращению экстрафузальныхволокон скелетных мышц и интрафузальныхволокон мышечных веретен. Гамма-эфферентный механизм — это, пожалуй, самый известный и наиболее изученный пример центрального контроля рецепторной функции. Волокна этого эфферентного пути идут к мышечным веретенам прямо от спинного мозга и проводят импульсы от продолговатого мозга и даже от центров еще более высокого уровня. Эти эфферентные волокна названы гамма-волокнами из-за их размеров (класс волокон Ay). Тела этих нейронов, так же как а-нейроны эффекторных мышечных нервов, находятся в спинном мозгу. Функция гамма-эфферентного механизма состоит в приспособлении сокращения мышечных волокон в веретене к данному состоянию скелетной мышцы. Импульсация гамма-эфферентных волокон, как правило, снижает порог возбуждения аннулоспиральных рецепторов. Этот механизм играет важную роль в поддержании определенного положения тела. Гамма-эфферентный механизм составляет часть саморегулирующейся системы. Когда тело или какая-либо часть его принимает определенное положение, растяжению соответствующих мыши предшествует приход гамма-эфферентной импульсации, регулирующей частоту импульсов, порождаемых аннуло- спиральными рецепторами. Затем центрально управляемые моторные нервы вызывают сокращение соответствующих мышц, причем активность этих нервов контролируется рефлек- торно гамма-эфферентами. Таким образом, эта система включает один афферентный и два эфферентных пути (ее называют также гамма-петлей). 11.Реципрокная иннервация мышц-антагонистов  Реципрокная иннервация (от лат. reciprocus — возвращающийся, обратный, взаимный), сопряжённая иннервация, рефлекторный механизм координации двигательных актов, обеспечивающий согласованную деятельность мышц-антагонистов (например, одновременное сокращение группы сгибателей сустава и расслабление его разгибателей). Сущность Реципрокная иннервация заключается в том, что рефлекторное возбуждение в группе нервных клеток, иннервирующих определённые мышцы, сопровождается реципрокным, т. е. сопряжённым, торможением активности в других клетках, функционально связанных с антагонистами, что ведёт к их расслаблению. Т. о., центры мышц-антагонистов — сгибателей и разгибателей — находятся в противоположном состоянии при выполнении многих двигательных актов. Механизм Реципрокная иннервация обеспечивает возможность осуществления организмом координированных движений (ходьба, чесание, движения глаз, трудовые движения и многие др.). Реципрокная иннервация была впервые обнаружена в 1876 П. А. Спиро, учеником И. М. Сеченова, и детально проанализирована английским физиологом Ч. Шеррингтоном, который и ввёл этот термин. Как показали Н. Е. Введенский и А. А. Ухтомский, этот механизм не жестко фиксирован, а динамичен, вследствие чего мышцы, являющиеся антагонистами при совершении одних движений, при участии в других сокращаются одновременно, т. е. ведут себя как синергисты. Прямое исследование процессов возбуждения и торможения в одиночных нервных клетках, проводящееся с помощью микроэлектродной техники с 50-х гг. 20 в., позволило понять особенности механизма Реципрокная иннервация на клеточном уровне. Ведущую роль в формировании сопряжённых отношений между двигательными нейронами, иннервирующими мышцы-антагонисты, играют вставочные нейроны, выполняющие в нервной системе функцию релейных переключателей и интегрирующих элементов. Из учебника: при вызове сгибательного рефлекса конечности импульсы от рефлексогенной зоны (кожа) поступают через вставочные нейроны к мотонейронам центра мышц-сгибателей, а также одновременно — к центру-антагонисту (мышц-разгибателей), но с включением на пути тормозного нейрона, который образует тормозной синапс на нейронах центра-разгибателя. Мышцы-разгибатели поэтому не сокращаются и не препятствуют сгибанию конечности, что осуществляется с помощью прямого постсинаптического торможения. Реципрокные взаимоотношения между центрами представлены довольно широко. Так, при возбуждении центра глотания тормозится центр жевания, возбуждение центра вдоха тормозит центр выдоха, рефлекс глотания тормозит вдох. 12. Спинальные двигательные рефлексы, их классификация. Рефлексы спинного мозга реализуются с использованием строго определенных его сегментов. КЛАССИФИКАЦИЯ СПИНАЛЬНЫХ РЕФЛЕКСОВ: • По количеству синапсов: –моносинаптические –полисинаптические • По количеству сегментов: –простые (1-2 сегмента) –сложные (более 2 сегментов) • По локализации рецепторов: –собственные –сопряженные • По характеру движений: –тонические –фазные –ритмические КЛАССИФИКАЦИЯ СПИНАЛЬНЫХ РЕФЛЕКСОВ Спинной мозг выполняет рефлекторную соматическую и рефлекторную вегетативную функции. Соматические спинальные рефлексы можно объединить в две группы по следующим признакам: - по рецепторам: а) проприоцептивные, б) висцероцептивные, в) кожные рефлексы. - по органам: а) рефлексы конечностей;б) брюшные рефлексы; в) рефлексы с органов малого таза (яичковый и анальный рефлексы). А) рефлексы конечностей: •Сгибательные фазные (локтевой рефлекс, Ахиллов, подошвенный рефлекс, акт ходьбы); тонические (поддержание позы). • разгибательные фазные (коленный рефлекс, локтевой разгибательный рефлекс, ходьба) тонические (поддержание позы стоя) • ритмические (чесательный и шагательный рефлексы) Б) Брюшные рефлексы проявляются при штриховом раздражении кожи живота. Для вызова верхнего брюшного рефлекса раздражение наносят параллельно нижним ребрам непосредственно под ними (Th8 –Th9). Средний брюшной рефлекс вызывают раздражением на уровне пупка (горизонтально), Th9 – Th10.Для получения нижнего брюшного рефлекса раздражение наносят параллельно паховой складке (рядом с ней), дуга рефлекса замыкается на уровне Th11 – Th12. в) рефлексы с органов малого тазаКремастерный (яичковый) рефлекс заключается в сокращении m. сremaster и поднимании мошонки в ответ на штриховое раздражение верхней внутренней поверхности кожи бедра (кожный рефлекс), это также защитный рефлекс. Его дуга замыкается на уровне L1 – L2 Анальный рефлекс выражается в сокращении наружного сфинктера прямой кишки в ответ на штриховое раздражение или укол кожи вблизи заднего прохода, дуга рефлекса замыкается на уровне S2 – S5 Вегетативные рефлексы спинного мозга осуществляются в ответ на раздражение внутренних органов и заканчиваются сокращением гладкой мускулатуры этих органов. Вегетативные рефлексы имеют в спинном мозге свои центры, которые обеспечивают иннервацию сердца, почек, мочевого пузыря и т.д. 135 стр учебника 13.Двигательные рефлексы спинного мозга в условиях их связи с вышестоящими отделами головного мозга. Перерезка или травма спинного мозга вызывает явление, получившее название спинального шока. Спинальный шок выражается в резком падении возбудимости и угнетении деятельности всех рефлекторных центров спинного мозга, расположенныхниже места перерезки. Деятельность центров, расположенных выше перерезки,сохраняется.  14.Роль мозга в регуляции фазных движений и мышечного тонуса. Продолговатый мозг и мост вместе с мозжечком входят в состав заднего мозга. В продолговатом мозге расположены ядра следующих пар черепно-мозговых нервов: 1 Двигательное ядро подъязычного нерва (пара XII) – иннервирует мышцы языка. 2 Двигательное ядро добавочного нерва (пара XI)– иннервирует голосовые мышцы, трапецевидныемышцы (наклоны головы вперед и вбок, пожимание плечами, их поднимание); 3 Группа ядер блуждающего нерва (пара X) – имеет 3 ядра: вегетативное иннервирует внутренние органы; чувствительное получает информацию от них и двигательное обеспечивает последовательность сокращения мышц глотки, гортани при глотании; 4 Языкоглоточный нерв (пара IX) – его ядро образовано 3 частями – двигательной, чувствительной и вегетативной. Двигательная часть участвует в иннервации мышц глотки и полости рта, чувствительная – получает информацию от рецепторов вкуса задней трети языка; 5 Преддверно-улитковый нерв (пара VIII) – состоит из улитковой и преддверной частей. Расположен на границе продолговатого мозга и моста. Преддверная часть формирует связи с вестибулярными ядрами моста (медиальным –Швальбе, латеральным – Дейтерса и верхним –Бехтерева). Функции продолговатого мозга. •Проводниковая. Через продолговатый мозг проходят все восходящие и нисходящие пути спинного мозга. Заканчиваются пути из коры большого мозга. •Рефлекторная. Многочисленные рефлексы продолговатого мозга условно делят на жизненно важные (дыхательные и сосудодвигательные центры) и не жизненно важные, защитные рефлексы (рвоты, чихания, кашля, слёзоотделения, смыкания век, жевания, глотания). В состав моста входят: 1 ядра лицевого нерва (пара VII), смешанный нерв, эфферентные волокна иннервируют мимические мышцы; 2 ядра отводящего нерва (пара VI), моторные волокна иннервируют мышцы, двигающие глазное яблоко, чувствительные – получают входы от проприорецепторов этих мышц; 3 тройничный нерв (пара V), обладает чувствительными и двигательными ядрами, последние иннервируют жевательные мышцы и мышцы неба. Функции моста •Проводящая функция моста. Здесь проходят, идущиеиз коры большого мозга пирамидные пути.  15.Рефлекторная деятельность среднего мозга. Ориентировочные рефлексы. Морфофункциональная организация. Средний мозг представлен четверохолмием и ножками мозга.Наиболее крупными ядрами среднего мозга являются: • красное ядро •черное вещество •ядра черепных (глазодвигательного и блокового) нервов •ядра ретикулярной формации. Сенсорные функции. Реализуются за счет поступления в него зрительной, слуховой информации. Проводниковая функция заключается в том, что через средний мозг проходят все восходящие пути к вышележащим таламусу (медиальная петля, спииноталамический путь), большому мозгу и мозжечку. Нисходящие пути идут через средний мозг к продолговатому и СМ. Это пирамидный путь, корково-мостовые волокна, руброретикуло- спинальный путь. Двигательная функция. Реализуется за счет ядра блокового нерва, ядер глазодвигательного нерва, красного ядра, черного вещества. • Ядро блокового (IV) нерва, иннервирует верхнюю косую мышцу глаза • Глазодвигательный (III) нерв иннервирует верхнюю, нижнюю и внутреннюю прямые мышцынижнюю косую мышцу и мышцу, поднимающую веко •расположенное в заднем мозге ядро отводящего (VI) нерва, иннервирует наружную прямую мышцу глаза. С участием этих ядер осуществляются поворот глаза в любом направлении, аккомодация глаза, фиксация взгляда на близких предметах путем сведения зрительных осей, зрачковый рефлекс (расширение зрачков в темноте и сужение их на свету). Рефлекторные функции. Функционально самостоятельными структурами среднего мозга являются бугры четверохолмия. Верхние из них являются первичными подкорковыми центрами зрительного анализатора, нижние — слухового. Основная функция бугров четверохолмия — организация реакции настораживания и так называемых старт-рефлексов на внезапные, еще не распознанные, зрительные или звуковые сигналы.Активация среднего мозга в этих случаях через гипоталамус приводит к повышению тонуса мышц, учащению сокращений сердца; происходит подготовка к избеганию, к оборонительной реакции. В ответ на внезапное раздражение происходит поворот головы и глаз в сторону раздражителя, а у животных—настораживания ушей. У человека возникает вздрагивание, иногда вскакивание на ноги, вскрикивание, максимально быстрое удаление от раздражителя, подчас бегство. Этот рефлекс необходим для подготовки организма к своевременной реакции на любое новое воздействие. Он сопровождается усилением тонуса мыщц-сгибателей (подготовка к двигательной реакции) и изменениями вегетативных функций (дыхание, сердцебиения). 16.Децеребрационная ригидность, механизм её возникновения. Децеребрационная ригидность — повышение тонуса всех мышц, чаще с резким преобладанием тонуса мышц-разгибателей в результате нарушения связей и разобщения головного мозга и мозгового ствола на уровне среднего мозга. ► Вызывается тем, что убираются стимулирующие влияния красного ядра на мотонейронымышц - сгибателей и остаются стимулирующие влияния ядра Дейтерса на мотонейроны мышц разгибателей после перерезки ствола мозга между средним и продолговатым мозгом (ниже красного ядра) ► Красное Ядро и Ретикулярная Формация (через рубро-спинальный и ретикулоспинальный латеральный пути) стимулируют сгибатели и тормозят разгибатели ► Ядро Дейтерса и Ретикулярная Формация (через вестибулоспинальный и ретикулоспинальный медиальный пути), наоборот, стимулируют мотонейроны разгибателей и тормозят мотонейронысгибателей Причины ригидности • опухоли и абсцессы мозга и мозжечка, внутримозговые гематомы. • отек и набухание мозга при обширном инфаркте мозга (обычно в первые 6 дней послеинсульта), ушибе мозга, менингите именингоэнцефалите, при токсических энцефалопатиях, почечной коме и др. 17.Статические и статокинетические рефлексы. Статические познотонические рефлексы. Обеспечивают поддержание естественной позы животного. Они осуществляются через продолговатый мозг с участием спинного: •рефлекс с вестибулярного аппарата на мышцы разгибатели конечностей. Обеспечивает их высокий тонус, сохранение положения позы животного теменем и спиной вверх; •рефлекс с вестибулярного аппарата на мышцы сгибатели конечностей. Возникает при положении животного теменем и спиной вниз и проявляется в повышении тонуса мышц сгибателей конечностей; •рефлекс с рецепторов мышц шеи на мышцы сгибатели задних конечностей и мышцы разгибатели передних конечностей. Проявляется при запрокидывании головы в выпрямлении передних конечностей и сгибании задних; •рефлекс с рецепторов мышц шеи на мышцы разгибатели задних конечностей и сгибатели передних конечностей. Проявляется в сгибании передних и разгибании задних конечностей при наклоне головы и шеи; Если над головой кошки держать кусок мяса, то она поднимает голову, при этом передние конечности разгибаются, задние сгибаются и животное садится, принимая позу, удобную для прыжка (А). Если же перед головой кошки поставить блюдце с молоком, она нагибает голову вниз, что вызывает сгибание передних и небольшое разгибание задних лап; в результате такого перераспределения тонуса кошка может начать лакать молоко (Б).Вестибулярные тонические рефлексы, зависящие от положения головы, у нормальной кошки. •рефлекс с рецепторов мышц шеи на мышцы разгибатели конечности одной стороны и мышцы сгибатели противоположной стороны. Проявляется при вращении в разгибании конечностей той стороны тела, в которую поворачивается голова, и в сгибании конечностей противоположной стороны. Выпрямительные рефлексы. Обеспечивают возвращение головы и тела из неестественного положения в естественное. Осуществляются через средний мозг: •рефлекс с рецепторов вестибулярного аппарата на мышцы головы возникает при положении головы и туловища на боку. Проявляется в перераспределении тонуса мышц головы и возврате головы в естественное положение; •рефлекс с тактильных рецепторов кожи при положении животного лежа на боку на мышцы головы. Обеспечивает возврат головы в естественное положение; •рефлекс с рецепторов мышц шеи, возникающий при изменении положения шеи, на мышцы туловища.Обеспечивает перевод туловища в положение, соответствующее положению шеи, за счет перераспределения тонуса мышц; •рефлекс с рецепторов кожи туловища, возникающий при положении животного на боку, на мышцы туловища. Обеспечивает за счет перераспределения тонуса мышц возврат туловища в естественное положение, соответствующее положению головы и шеи. Статокинетические рефлексы. Проявляются при движении животного, изменении положения отдельных частей тела, когда происходит перераспределение тонуса мышц глаз, туловища и конечностей, что обеспечивает устойчивое положение глаз, головы и тела в пространстве: •рефлекс с рецепторов мышц одной конечности на мышцы других. Возникает при ходьбе животного, когда при сгибании одной конечности повышается тонус мышц разгибателей других трех конечностей; •рефлекс «нистагм головы» возникает при вращательных движениях. Проявляется в движении головы в противоположную сторону вращения туловища, а затем в скачкообразномперемещении головы в положение, соответствующее положению туловища; • рефлекс «нистагм глаз» возникает при вращательных движениях; Проявляется в движении глаз в противоположную сторону вращения головы и туловища, а затем в скачкообразном перемещении глаз в положение, соответствующее положению туловища; • лифтовый рефлекс возникает при линейном ускорении движения вверх и вниз; в первом случае повышается тонус мышц разгибателей, во втором — тонус мышц сгибателей. 18.Базальные ядра, роль, проявления поражений.  БАЗАЛЬНЫЕ ЯДРА Скорлупа + Хвостатое Ядро = Полосатое тело (стриатум) Бледный шар (наружный сегмент) Бледный шар (внутренний сегмент) Субталамическое ядро Черная субстанция Миндалина Афференты к стриатуму: • от всех областей коры • от таламуса • от черной субстанции среднего мозга через дофаминэргический путь; Эфференты от стриатума: • в бледный шар; • к черной субстанции; • через таламус в двигательную кору Функции базальных ядер 1 Центры координации сочетанных двигательных актов 2 Центры контроля координации тонуса мышц и произвольных движений 3 Центры сложных безусловных рефлексов и инстинктов 4 Центры торможения агрессивных реакций 5 Участие в механизмах сна Расстройства и болезни, связанные с базальными ганглиями Симптомы поражения полосатого тела: Хорея (хореический гиперкинез, также известная как «виттова пляска» или «пляска святого Витта») — синдром, характеризующийся беспорядочными, отрывистыми, нерегулярными движениями, сходными с нормальными мимическими движениями и жестами, но различные с ними по амплитуде и интенсивности, то есть более вычурные и гротескные, часто напоминающие танец. Торсионная дистония. Синдром, при котором происходит постоянное спазматическое сокращение мышц, затрагивающее как мышцу-агонист, так и противодействующую ей мышцу. Спазмы мышц часто непредсказуемы, они изменяют нормальное положение тела, могут носить хронический характер и вызывать значительное неудобство, боль и потерю трудоспособности. Появляются патологические позы – штопорообразные движения туловища или локальные спазмы (в виде кривошеи). Атетоз (от др. – греч. ἄθετος — неустойчивый) — гиперкинез (патологические непроизвольные движения), выражающийся медленной тонической судорогой конечностей, лица, туловища. Отличается непроизвольными медленными стереотипными, вычурными движенияминебольшого размаха в дистальных отделах конечностей, которые могут распространяться и на проксимальные отделы конечностей, а также мышцы головы, языка, лица (вытягивание губ, перекашивание рта, гримасничанье). Атетоз (медленные червеобразные, вычурные движения кистей рук, пальцев).  Гемибаллизм – быстрые беспорядочные сокращения мышц – размашистые движения конечностей, вращательные движения. Миоклония– внезапные непроизвольные сокращения одной или нескольких групп мышц, возникающие как при движениях, так и в состоянии покоя. Симптомы поражения бледного шара Болезнь Паркинсона — медленно прогрессирующее хроническое неврологическое заболевание, характерное для лиц старшей возрастной группы. Относится к дегенеративным заболеваниям экстрапирамидной моторной системы. Вызвано прогрессирующим разрушением и гибелью нейронов, вырабатывающих нейромедиатор дофамин, — прежде всего в чёрной субстанции, а также и в других отделах центральной нервной системы. Недостаточная выработка дофамина ведет к активирующему влиянию базальных ганглиев на кору головного мозга. Ведущими симптомами (иначе: основные или кардинальные симптомы) являются: •мышечная ригидность; •гипокинезия; •Тремор; •постуральная неустойчивость. Синдром Туретта (болезнь Туретта, синдром Жиля де ла Туретта) — генетически обусловленное расстройство центральной нервной системы, которое проявляется в детском возрасте и характеризуется множественными моторными тиками и как минимум одним вокальным или механическим тиком. Ранее синдром Туретта считался редким и странным синдромом, ассоциируемым с выкрикиванием нецензурных слов или социально неуместных и оскорбительных высказываний (копролалия). Значение базальных ядер ---участвуют в интеграции тонических рефлексов; ---являются одним из уровней системы регуляции движений, передают в основном тормозные влияния кмоторной коре и в ствол мозга; ---являются важнейшим связующим звеном между ассоциативными и моторными областями коры больших ---участвуют в создании программ целенаправленных движений. Их роль важна в переходе от замысла движений (фаза подготовки) к выбранной программе действия (фаза выполнения), а также в формировании необходимой для выполнения движения позы. 19.Функциональная и нейронная организация мозжечка. Мозжечок располагается у человека позади больших полушарий над продолговатым мозгом и мостом мозга. В эволюционном плане мозжечок представляет собой очень древнюю структуру. Анатомия мозжечка • средний отдел (червь); •примыкающие области (дольки старой коры или клочки) связанные с вестибулярными ядрами продолговатого мозга – вестибулоцеребеллум ; •полушария мозжечка, которые функционально принято разделять на две области: расположенную ближе к червю – спиноцеребеллум и латеральную-цереброцеребеллум. В полушариях мозжечка выделяют –трехслойную кору мозжечка и скопления нервных клеток – четыре пары ядер: зубчатого, пробковидного, шаровидного и кровельного. Нейронная организация коры мозжечка В ней выделяют три слоя: I – поверхностный, или молекулярный слой (дендритные разветвления грушевидных нейронов - клеток Пуркинье; корзинчатыеклетки, звездчатые клетки); II – ганглиозный слой (клетки Пуркинье); III – гранулярный слой (клетки-зерна, или гранулярные клетки; клетки Гольджи ).  20.Афферентные и эфферентные связи мозжечка с другими отделами мозга.   21.Роль мозжечка в координации деятельности экстрапирамидной и пирамидной систем. Вестибулоцеребеллум : его основная задача состоит в сохранении равновесия при стоянии и ходьбе, а также в управлении движениями глаз. Спиноцеребеллум контролирует правильность начинающихся движений ног и рук. Цереброцеребеллумучаствует в планировании движений. ---участвует в регуляции позы, мышечного тонуса и равновесия; ---осуществляет координацию целенаправленных движений с рефлексами поддержания позы; ---производит координацию быстрых целенаправленных движений, осуществляемых по команде из коры больших полушарий, таких как бег, прыжки, игра на фортепьяно и даже речь ; ---является хранилищем центральных двигательных программ. В нем хранятся программы сложных и автоматически выполняемых двигательных актов. Он также корректирует выполнение двигательных программ. 22.Проявления поражения мозжечка у животных и человека. триада Лючиани: а) атония, б) астазия, в) астения, триада Шарко: нистагм, интенционный тремор и скандированная речь.  Поражение мозжечка приводит к дистонии -неадекватному перераспределению тонуса мышц. При этом сразу же после повреждения мозжечка наблюдается гипертонус мышц разгибателей, который через некоторое время сменяется гипотонией - снижением тонуса мышц или атонией – полным отсутствием мышечного тонуса. Астазия - нарушение равновесия, качательныедвижения при стоянии; Дизартрия - расстройство артикуляции. Речь становится медленной, невыразительной, монотонной. Асинергия - нарушение содружественныхдвижений. Целостное движение состоит не из одновременных содружественных актов, а из последовательного ряда простых движений. Так, например, касание кончика носа мозжечковый больной осуществляет в три приема (сначала опускает руку, затем сгибает ее в локте и только после этого подносит палец к носу); Деэквилибрация - нарушение равновесия (выявляется проведением пробы Ромберга, спомощью которой проверяется способность удерживать равновесие при закрытых глазах, когда ноги поставлены пятками вместе, а руки вытянуты вперед); Атаксия - нарушение точности и координации движений Дисметрия - утрата соразмерности движений, что особенно наглядно проявляются при совершении целенаправленных движений, когда конечность либо не достигает цели, либо проносится мимо нее; Адиадохокинез - неспособность быстро и равномерно выполнять противоположные движения, например, быстро поворачивать руку то ладонью вверх, то ладонью вниз; 23.Функциональная и нейроннаяорганизация двигательных зон коры больших полушарий.Основные проявления нарушений деятельности моторной коры и кортикоспиральных трактов. Кора головного мозга - высший отдел центральной нервной системы, обеспечивающий функционирование организма как единого целого при его взаимодействии с окружающей средой.        Первичная моторная кора содержит гигантские пирамидные клетки Беца, их аксоны формируют кортикоспинальный и кортикобульбарный тракты. Кортико-спинальный путь (пирамидный) представляет собой единственную прямую связь головного мозга со спинным. При повреждении пирамидного тракта: 1 Исчезают или резко ограничиваются произвольные движения со снижением мышечной силы; 2 Спастическая гемиплегия (параличи); 3 Усиливаются глубокие миотатическиерефлексы, исчезают кожные, появляются патологические рефлексы; 4 Синкинезия – центральный паралич; 5 Сопротивление пассивным движениям по типу складного ножа (сопротивление только в начале движения); 6 Поза Вернике-Мана – повышен тонус сгибателей рук и разгибателей ног («рука просит, нога косит»). Своеобразные нарушения движений возникают при повреждениях премоторной коры, дополнительных двигательных областей коры и задних отделов теменной коры — тех областей коры, которые играют решающую роль в планировании всего комплекса движений, необходимых для достижения определенной цели в той или иной конкретной обстановке. Человек утрачивает способность воспроизводить (повторять) предъявляемые ему движения. Утрачивает ранее приобретенные двигательные навыки (навыки игры на музыкальных инструментах, шитья, вязания, пользования ножом и вилкой и т.п.). При разрушении двигательной зоны центральной извилины и лобной зоны впереди нее (4, 6, 8, 9 поля Бродмана) – нарушения двигательных функций: адинамия, парез, паралич (соответственно ослабление, резкое снижение, исчезновение движений). При разрушении зрительной зоны: 17 поля - возникает выпадение зрительных ощущений (корковая слепота); 18 поля Бродмана - страдают функции, связанные с распознаванием зрительного образа, нарушается восприятие письма; 19 поля Бродмана - возникают различные зрительные галлюцинации, страдает зрительная память и другие зрительные функции. При поражении слуховой зоны (височная область коры головного мозга): 42 поля - нарушается функция распознавания звуков. 22 поля - возникают слуховые галлюцинации, нарушение слуховых ориентировочных реакций, музыкальная глухота. 41 поля - корковая глухота. При поражении речедвигательного центра Брока мышцы речедвигательного аппарата интактны, но человек не способен говорить, как ребенок первых месяцев жизни. Это состояние называется моторной афазией;При поражении центра Вернике возникает сенсорная афазия - человек не понимает устную речь (как чужую, так и свою). Из-за непониманиясобственной речевой продукции речь больногоприобретает характер «словесного салата», т.е. набора не связанных между собой слов и звуков. Агнозии - это расстройства узнавания и познавания, отражающие нарушения различных видов восприятия (формы предмета, символов, пространственных отношений, звуков речи и т.д.), возникающие при поражении коры больших полушарий головного мозга. Апраксии - расстройства праксиса, которые характеризуются утратой навыков, выработанных в процессе индивидуального опыта, сложных целенаправленных действий (бытовых, производственных, символической жестикуляции) без выраженных признаков центрального пареза или нарушений координации движений. Вывод: Двигательная кора за счет согласованной деятельности пирамидной, экстрапирамидной и мозжечковой систем обеспечивает целенаправленные произвольные движения, двигательные навыки, координацию и программирование движения. |