Физиология. Контрольные вопросы занятие 5. Биоэлектрические явления в возбудимых тканях. Методы исследования возбудимых тканей
 Скачать 298.29 Kb. Скачать 298.29 Kb.
|

|
КОНТРОЛЬНЫЕ ВОПРОСЫ Занятие №5. Биоэлектрические явления в возбудимых тканях. Методы исследования возбудимых тканей 1) Возбудимые ткани. Общие свойства возбудимых тканей. Возбудимость и возбуждение. Специфические и неспецифические признаки возбуждения. Критерии оценки возбудимости. Классификация раздражителей. Биоэлектрические процессы являются основой быстрой и пространственно специфической реакции организма на изменение внешней и внутренней среды. Возбудимые ткани – это ткани, способные отвечать на действие раздражителя активной физиологической реакцией; к возбудимым тканям относят те, в которых четко прослеживается переход из состояния покоя в состояние возбуждения (нервная, мышечная, железистая). Общие свойства возбудимых тканей: возбудимость; проводимость; лабильность; рефрактерность; сократимость (мышечная ткань); секреция (железистая ткань). Возбудимость – способность клеток реагировать определенным образом на действие раздражителя. Возбуждение – активный физиологический процесс, сопровождающийся изменением функционального состояния клетки, вызванный действием внешних или внутренних факторов.   Признаки возбуждения
Раздражитель – фактор внешней среды, воздействующий на биологический объект (возбудимую ткань).    Раздражители
2) Электрические явления в возбудимых тканях. История их открытия (Л. Гальвини, А. Вольта, Ч. Маттеучи). История изучения в данной области начинается с конца XVIII века (опыты Гальвани, Вольта и др.). Суть первого опыта Гальвани заключается в сокращении икроножной мышцы лягушки при действии особым (Fe/Cu) пинцетом на седалищный нерв, а второго опыта Гальвани – в сокращении при набрасывании на нее седалищного нерва на поврежденный и неповрежденный участок поверхности мышцы. 3) Современные представления о строении и функциях мембран, ионных каналов. Активный и пассивный транспорт ионов через мембраны. В отличие от изучения электрических процессов на макроуровне, расшифровка структуры мембран сравнительно молодое направление физиологии. Вначале была предложена модель «сэндвича», согласно которой основные компоненты располагались слоями, затем жидко-мозаично, т.е. в билипидном слое отдельно располагаются на поверхности (периферические) или насквозь белковые молекулы (интегральные). Конев С.В. и др. (1970) выдвинули твердо-каркасную жидко-мозаичную модель: белковый компонент мембраны образует скелет мембраны (твердоупругий белковый каркас), ячейки которого заполнены липидным бислоем. Цитоскелет регулирует конформацию, подвижность, функциональную активность элементарных белков типа рецепторов, ферментов, транспортных систем или ионных каналов, с другой стороны, он придает мембране ряд интегральных свойств (непрерывность, прочность, эластичность). Функции клеточной мембраны: защитная; барьерная; рецепторная; транспортная; ферментативная.   Классификация типов транспорта низкомолекулярных веществ без изменения морфологической структуры мембраны 
А. Пассивный транспорт – перенос веществ через биомембрану по градиенту (концентрационный, осмотический, гидродинамический и т.д.) и без расхода энергии. Простую пассивную диффузию описывает закон Фика. Облегчённая диффузия – это вид переноса ионов через биологические мембраны, который осуществляется по градиенту концентрации с помощью переносчика. Ионный канал – это несколько субъединиц (интегральных мембранных белков, содержащих трансмембранные сегменты, каждый из которых имеет – спиральную конфигурацию), обеспечивающих перенос ионов через мембрану. Б. Активный транспорт – перенос веществ через биомембрану против градиента и с расходом энергии. У человека 30-40% всей энергии, образующейся в ходе метаболических реакций, расходуются на активный транспорт. В почках 70-80% потребляемого кислорода идет на активный транспорт. В 1957 г. Я. Скоу обнаружил фермент, гидролизирующий АТФ при добавлении ионов Na+ и К+ (Na+/К+-АТФ-аза) – этот фермент состоит из двух полипептидных цепей. Na+/K+-насос – энергозависимый механизм, обеспечивающий выведение из клетки ионов натрия и введение в нее ионов калия. При гидролизе одной молекулы АТФ этим ферментом происходит поступление двух ионов К+ внутрь клетки и выведение трёх ионов Na+ из клетки. 4) Мембранный потенциал покоя, его происхождение и механизм поддержания (селективная проницаемость, Na+/K+-насос и др.). Мембранный потенциал – разность потенциалов между цитоплазмой и окружающей средой. Для этого вводят один электрод внутрь клетки, а другой располагают снаружи. Впервые его измерение было выполнено в 1939 г. А. Ходинским, А. Хаксли на гигантском аксоне кальмара. Различают два вида мембранных потенциалов: потенциал покоя и потенциал действия. А. Потенциал покоя – это мембранный потенциал клетки, находящейся в невозбуждённом состоянии. Величина потенциал покоя равна -60 – -90 мВ для волокон поперечно-полосатой мускулатуры, -30 – -50 мВ – для волокон гладкой мышечной ткани, -60 – -70 мВ – для нервных клеток. Внутри клетки содержится избыточный отрицательный заряд. В его возникновении ключевая роль принадлежит мембране, создающей неравномерное распределение ионов внутри и вне клетки. Мембрана обладает селективной проницаемостью для разных ионов. В соответствии с мембранно-ионной теорией возбуждения генез биопотенциалов в клетках возбудимых тканей обусловлен: градиентом концентраций ионов Na+, K+, Ca2+, Cl- внутри и вне клетки; селективной проницаемостью для них клеточной мембраны; функционирование Na+/K+-насосом, обеспечивающим поддержание концентрационной разницы. Ионы К+ легко диффундируют через мембрану посредством существующих механизмов переноса. Мембрана обладает избирательной проницаемостью для ионов К+ и практически непроницаема для других ионов, и прежде всего Na+. Величину мембранного потенциала можно рассчитать в соответствии с уравнением Нернста: а) только для К+: 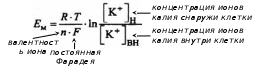 б) для нескольких ионов: 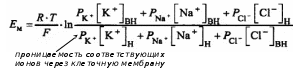 Ионы К+ диффундируют через мембрану, располагаются снаружи, создавая положительный заряд, соответствующий по величине отрицательному заряду внутри. Выход положительных ионов К+ создает электрическое поле, которое мешает выходу других ионов К+ за счёт электроосмотического давления. Однако мембрана не полностью непроницаема для ионов Na+. Эти ионы входят внутри клетки пассивно. Непрерывное поступление Na+ в клетку и выход из нее К+ должно было бы нарушить существующее равновесие ионов. Внутриклеточные концентрации Na+ должны были бы возрастать, а К+, наоборот, уменьшаться, что, в конечном итоге, вело бы к снижению потенциала покоя. В обычных условиях этого не происходит. Существует дополнительный механизм, обеспечивающий поток ионов против градиента концентраций, требующий затрат энергии. Это доказывается опытами с радиоактивным изотопами Nа+ (его вводят внутрь клетки, а затем в процессе жизнедеятельности регистрируют вне). При охлаждении нервной клетки на 0,5 °С выход Na+ снижается в 10 раз, а при нормализации температуры его поток возрастает (эта температурная зависимость не может наблюдаться за счет пассивной диффузии). В опытах с динитрофенолом (блокатором метаболических процессов) было показано снижение концентрации ионов Na+. 5) Потенциал действия и его фазы согласно мембранно-ионной теории возбуждения. Критический уровень деполяризации. В настоящее время процессы возбуждения описываются, исходя из представлений мембранно-ионной гипотезы Бернштейна, согласно которой изменение мембранного потенциала под действием раздражителей обусловлено изменением специфической ионной проницаемости. Б. Потенциал действия – это кратковременное изменение мембранного потенциала клетки при её возбуждении. Продолжительность потенциала действия может изменяться в широком временном диапазоне (от 1 мсек – для нервных клеток, до 10 мсек – (волокна скелетной мускулатуры) и даже 100-300 мсек для кардиомиоцитов и более – для гладкой мускулатуры). Потенциал действия имеет несколько фаз: фаза локального ответа; фаза деполяризация; фаза реполяризация; фаза следовых потенциалов (следовая деполяризация и гиперполяризация), положительную часть называют овершутом. Во время фазы деполяризации формируется быстрый поток Na+, его проницаемость резко возрастает. Накопление Na+ в клетки приводит к уменьшению величины отрицательного заряда и приобретение некоторого положительного заряда. Затем поступление Na+ прекращается и наблюдается выход К+ из клетки, что формирует фазу реполяризации. В основе быстрого поступления Na+ в клетку лежит активация Na+-каналов, которые представляют собой специальные потенциалзависимые структуры, образованные протеинами. Они имеют определенный диаметр примерно 0,3 нм, располагаются примерно на расстоянии 140 нм. Эти каналы являются селективными. Внутри канала имеется особый воротный механизм, который открывается в процессе возбуждения. Для К+ существуют аналогичные специальные К+-каналы, которые имеют также специальный воротный механизм, открывающийся при деполяризации. Генерация потенциала действия не требует затрат энергии! При действии слабых раздражителей в мембране происходит пассивная электротоническая деполяризация. При увеличении силы раздражителя на мембране деполяризация возрастает и длительное время сохраняется локальный ответ. В мембране при этом изменяется ионная проницаемость, изменяется возбудимость (растет). I. Локальный ответ – возбуждение, возникающее при действии на возбудимую ткань подпороговых раздражителей. II. Деполяризациия – это изменение мембранного потенциала в менее электроотрицательном направлении. Возникновение фазы деполяризации обусловлено открытием Na+-каналов и вхождением ионов Na+ в клетку. Критический уровень деполяризации – это уровень мембранного потенциала, при котором открываются все потенциал-зависимые натриевые каналы и начинается деполяризация. Порог деполяризации есть разница между потенциалом покоя и величиной критического уровня деполяризации. III. Реполяризация – восстановление исходного уровня мембранного потенциала. Возникновение фазы реполяризации обусловлено инактивацией Na+-каналов, открытием К+-каналов и выходом этих ионов из клетки. IV. Фаза следовых потенциалов – период потенциала действия, возникающий после реполяризации, в котором выделяют деполяризационную и гиперполяризационную части. Овершут – это фрагмент потенциала действия, во время которого внутреннее содержимое клетки приобретает положительный заряд по отношению к окружающей среде. Гиперполяризация – это изменение мембранного потенциала в более электроотрицательном направлении. 6) Соотношение фаз возбудимости с фазами потенциала действия. Рефрактерность и её причины. Особенности местного и распространяющегося возбуждения. П 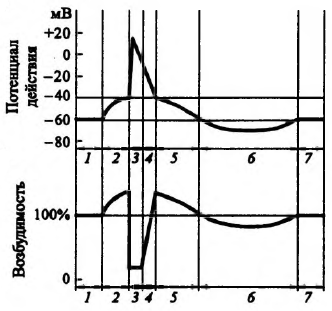 отенциал действия: отенциал действия:1, 7 – потенциал покоя; 2 – локальный ответ; 3 – деполяризация и начальная реполяризация; 4 – окончательная реполяризация; 5 – следовая деполяризация; 6 – следовая гиперполяризация. Возбудимость: 1, 7 – исходная возбудимость; 2 – супернормальная возбудимость; 3 – абсолютная рефрактерность; 4 – относительная рефрактерность; 5 – супернормальная возбудимость; 6 – субнормальная возбудимость. Рефрактерность – состояние невозбудимости клетки во время её возбуждения: абсолютная (полная) и относительная (неполная). Аккомодация – уменьшение возбудимости ткани при действии медленно нарастающего по силе раздражителя. В её основе лежат процессы инактивации натриевой и повышения калиевой проницаемости мембраны. Сравнительная характеристика локального потенциала и потенциала действия
7) Законы раздражения возбудимых тканей (закон «всё или ничего», значение силы, длительности, крутизны нарастания раздражения). Хронаксиметрия. 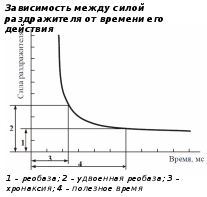 Суть закона «всё или ничего» состоит в том, что при деполяризации клетки в данных условиях амплитуда всех потенциалов, которые она генерирует, будет одинакова - объясняется это уравнением Нерста для случая открытых натриевых каналов. Изучалась ответная реакция в зависимости от силы и длительности раздражения. Зависимость пороговой силы раздражителя от его длительности исследовалась Гоорвег (1892), Вейс (1909), Лапик (1909). Была получена кривая «сила-время». Существует зависимость между силой и длительностью действия раздражителя и ответной реакцией на его действие. Для исследования данной закономерности используют метод хронаксиметрии, согласно которому определяют хронаксию и реобазу. Реобаза – минимальная сила тока, которая способна возбудить данную ткань. Хронаксия – минимальное время действия тока, при котором ток силой в две реобазы способен возбудить данную ткань. Закон «силы» – закономерность, согласно которой при увеличении силы раздражителя ответная реакция возбуждения растёт. 8) *Действие постоянного тока на возбудимые ткани. Полярный закон раздражения. Электротонические явления в тканях, их значение в проведении возбуждения. Катодическая депрессия, анодическая экзальтация. Физиологический электротон – это изменение возбудимости клеток или тканей под действием постоянного электрического тока. Катэлектротон – это изменение возбудимости клеток или тканей под действием постоянного тока под катодом. Анэлектротон – это изменение возбудимости клеток или тканей под действием постоянного тока под анодом. Катодическая депрессия – это снижение возбудимости под катодом при длительном действии постоянного тока подпороговой величины. Анодическая экзальтация – это повышение возбудимости под анодом при длительном действии постоянного тока подпороговой величины. |