Лекции биология. Курс лекций для студентов, обучающихся на русском языке Рязань 2008
 Скачать 7.22 Mb. Скачать 7.22 Mb.
|

|
Функционально-генетическая классификация генов В настоящее время ген рассматривается как единица функционирования наследственного материала. Ген – это участок молекулы ДНК, ответственный за синтез одного полипептида. Различают три вида генов: - структурные; - модуляторы; - регуляторы. Структурные гены несут информацию об аминокислотах в белках, ферментах, а также о последовательности нуклеотидов в молекулах р-РНК и т-РНК. Гены-модуляторы влияют на функционирование структурных генов, могут смещать в ту или иную сторону процесс развития признака. Они подразделяются на: 1)ингибиторы или супрессоры (эпистатичные гены), 2)интенсификаторы – могут повышать способность структурных генов к мутациям, 3)модификаторы – могут влиять на структурные гены по типу комлементарности. Гены-регуляторы контролируют синтез регуляторных белков, а также время включения различных структурных генов в процессе индивидуального развития. Свойства генов: 1. Дискретность действия – каждый ген действует как самостоятельная единица наследственности. 2. Стабильность – при отсутствии мутаций ген передается в ряду поколений в неизмененном виде. 3. Специфичность действия – каждый ген влияет на развитие своего определенного признака. 4. Плейотропия – способность одного гена обеспечивать развитие одновременно нескольких признаков (синдром Марфана). 5. Присутствие в виде двух у диплоидных и большего числа аллелей у полиплоидных организмов. 6. Действие гена дозировано, при изменении числа доз гена в организме изменяется признак (например, при болезни Дауна происходит увеличение до трех доз генов 21 хромосомы) Функциональная активность генов или экспрессия генов У прокариот она осуществляется в два этапа: транскрипция и трансляция.У эукариот есть еще стадия процессинга. Экспрессия генов заключается в синтезе на молекуле ДНК молекулы и-РНК, комлементарной ей (или транскрипции – переписывание, считывание биологической информации) и дальнейшее ее использование для синтеза белка. Единицей транскрипции в ДНК является транскриптон, превышающий по размерам структурные гены. Транскриптон в клетках эукариот состоит из неинформативной (акцепторной) и информативной зоны. Неинформативная зона начинается геном-промотором (участок из 80 нуклеотидов), к которому присоединяется фермент РНК-полимераза, катализирующая процесс считывания. У прокариот один вид РНК-полимеразы, у эукариот три. За геном промотором находятся гены-операторы, которые связывают регуляторные белки (белки, включающие и прекращающие транскрипцию). Информативная зона состоит из структурных генов, располагающихся за генами операторами. Структурные гены эукариот разделены спейсерами – участками ДНК, не несущими информации. Кроме того, в составе самих структурных генов есть информационные участки – экзоны и неинформационные – интроны. У пркариот спейсеров, экзонов и интронов нет. В каждой фазе жизненного цикла в клетке транскрибируется только 10% структурных генов, а остальные гены не активны, но часть из них может включаться в других фазах жизненного цикла. В результате транскрипции у прокариот сразу образуется м-РНК (зрелая РНК) и сразу же начинается процесс трансляции. У эукариот транскрибируется большая молекула и-РНК, содержащая все неинформативные участки. Она называется РНК-предшественница или пре-РНК. Поэтому за транскрипцией наступает процессинг, в результате, которого разрушаются все неинформативные участки: акцепторная зона, спейсеры и интроны, а оставшиеся экзоны сшиваются (сплайсинг). На этапе процессинга к начальному отрезку образовавшейся РНК присоединяется последовательность нуклеотидов, называемая колпачком, к концевому – последовательность остатков аденина – поли- A. В процессинге происходит модификация нуклеотидов в РНК, например, их метилирование, гидрирование. И только после этих превращений образуется зрелая м-РНК, которая начинается вводной последовательностью, называемой лидером, и заканчивается концевой последовательностью – трейлером. Лидер – вводная последовательность нуклеотидов, комплементарная последовательности в молекуле р-РНК малой субъединице рибосом, которая обеспечивает прикрепление и-РНК к малой субъединице. Трейлер включает нонсенс-кодон и поли-А последовательность. Кодовым элементом м-РНК является триплет нуклеотидов, называемый кодоном. Каждому кодону соответствует определенная аминокислота. Первичной структурой РНК является, как и в ДНК, линейная цепь полинуклеотидов, в которой мононуклеотиды соединены 3`,5`-фосфоэфирными связями. Вторичная структура РНК – изогнутая цепь, а третичная представляет собой нить, намотанную на катушку, роль катушки играет особый транспортный белок – информатор. Образующаяся м-РНК идет в цитоплазму к месту синтеза полипептида (белка), т.е. к рибосомам. На рибосомах осуществляется процесс трансляции. Трансляция – это механизм, с помощью которого последовательность нуклеотидов в молекулах м-РНК переводится в специфическую последовательность аминокислот в полипептидной цепи. Трансляция складывается из трех стадий: 1. Инициация – начало синтеза полипептида. 2. Элонгация – удлинение полипептида. 3. Терминация – окончание синтеза полипептида. На этапе инициации меньшая субъединица рибосомы узнает стартовый кодон АУГ м-РНК и прикрепляется к ней. АУГ занимает первую позицию. После этого присоединяется большая субъединица рибосомы и в ней начинается собственно синтез белка. К большой субъединице подходит т-РНК с аминокислотой. 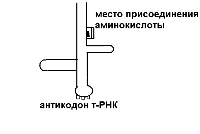 В молекуле т-РНК одна ее часть присоединяет аминокислоту, а другая – антикодон, спаривается с колоном м-РНК, определяющим эту аминокислоту. 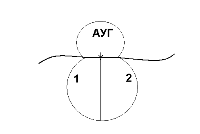 В большой субъединице есть: 1 – аминоацильный участок 2 – пептидильный участок В т-РНК, антикодон которой комплементарен кодону АУГ м-РНК приносит аминокислоту метианин и останавливается в аминоацильном участке. Стадия инициации контролируется факторами инициации. У прокариот их 3, у эукариот 6-7. Вторая стадия – элонгации начинается с перемещения т-РНК с метианином в пептидильный участок, а на ее место приходит другая т -РНК с новой аминокислотой. Между двумя аминокислотами образуется пептидная связь. Тем временем рибосома продвигается вдоль м-РНК, на рибосоме оказывается новый кодон, к которому вскоре присоединяется своим антикодоном соответствующая т-РНК. Все это повторяется многократно, до тех пор, пока рибосома не дойдет до одного из терминальных кодонов (УАА, УАГ, УГА). Этим кодонам нет антикодона, нет аминокислоты. Наступает фаза терминации. К одной молекуле м-РНК прикрепляется обычно много рибосом и образуются полисомы. В процессе трансляции наряду с факторами инициации и элонгации важное значение имеет фермент, связывающий т-РНК с аминоацильным участком – аминоацил т-РНКаза. Регуляция экспрессии генов у про- и эукариот У прокариот она осуществляется на уровне промотора, оператора и трансляции. У эукариот регуляция происходит на всех этапах экспрессии: транскрипции, процессинга и трансляции. Подробнее остановимся на регуляции экспрессии генов у бактерий. Регуляция экспрессии генов у прокариот 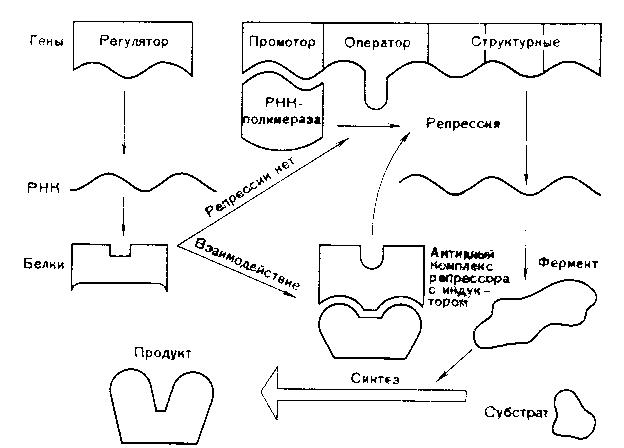 Схема регуляции транскрипции структурных генов прокариотической клетки по типу репрессии 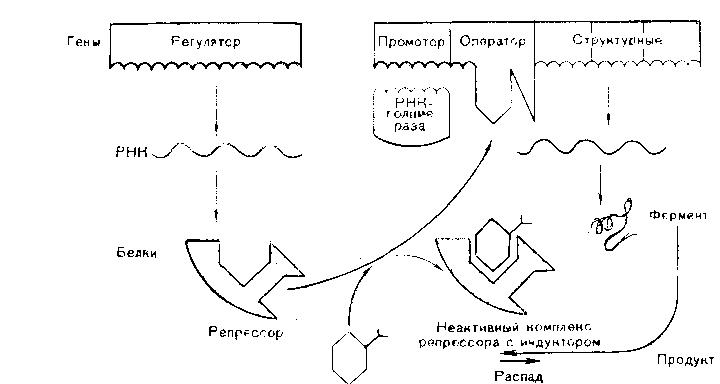 Схема регуляции транскрипции структурных генов прокариотической клетки по типу индукции В 1961 году Жакоб и Моно установили, что у бактерий под контролем гена-регулятора синтезируется белок-репрессор, который регулирует активность других генов. Белок-репрессор связывается с геном-оператором и блокирует его, вследствие чего транскрипция (считывание информации) становится невозможной и клетка не может синтезировать соответствующие белки. Но если в клетку проникает какой-либо субстрат, для расщепления которого необходимо синтезировать ферменты, то происходит следующее: субстрат присоединяется к репрессору и лишает его способности блокировать ген-оператор. К ДНК присоединяются РНК-синтезирующие ферменты и начинается транскрипция генов. Синтез белка-фермента прекращается, если субстрат полностью расщепляется, белок-репрессор освобождается и снова блокирует гены-операторы. Это пример регуляции по типу индукции. Также существует регуляция по типу репрессии: когда конечные продукты биохимической реакции, соединяясь с неактивным белком-репрессором, образуют комплекс, блокирующий работу гена-оператора. Антитерминация – заключается в том, что происходит игнорирование терминальных кодонов, процесс экспрессии продолжается. Регуляция у эукариот 1)Регуляция генной активности у эукариот намного сложнее, чем у бактерий. У эукариот она происходит не только на уровне клетки. Существуют системы регуляции организма как целого. Огромную роль в регуляции играют гормоны, но регулируют они процессы синтеза белков лишь в клетках-мишенях. Гормоны связываются с белками-рецепторами, расположенными в мембранах таких клеток и включают системы изменения структуры клеточных белков. Те, в свою очередь, могут влиять как на процессы транскрипции, так и процессы трансляции. Каждый гормон через систему посредников активирует свою группу генов. Так адреналин включает синтез ферментов, расщепляющих гликоген мышц до глюкозы, а инсулин влияет на образование гликогена из глюкозы в печени. 2)На стадии транскрипции белки-гистоны участвуют в процессах регуляции генной активности у эукариот. Непременное условие – это деконденсация участка, где происходит транскрипция. 3)Регуляция на уровне трансляции направлена на сохранение стабильности м-РНК, а эффективность трансляции осуществляется с помощью факторов инициации, элонгации, терминации. Лекция 12. Изменчивость План 1.Определение и формы изменчивости: модификационная, комбинативная, мутационная. 2.Мутагенные факторы. 3.Классификация мутаций. 4.Устойчивость и способы репарации генетического материала. 5.Закон гомологических рядов наследственной изменчивости Н.И.Вавилова. Определение и формы изменчивости Генетика изучает два основных свойства живых существ - наследственность и изменчивость. Изменчивость - свойство организмов приобретать новые признаки и особенности индивидуального развития под влиянием факторов среды. Изменчивость - один из важнейших факторов эволюции, обеспечивающих все многообразие живой природы. Различают два вида изменчивости: 1. Фенотипическую (ненаследственную, модификационную); 2. Генотипическую (наследственную): а) комбинативную; б) мутационную. Модификационная изменчивость – форма изменчивости, не связанная с изменением генотипа и вызванная влиянием факторов среды. Модификационная изменчивость имеет особенности: - не затрагивает наследственную основу организма и поэтому модификации не передаются по наследству, то есть от родителей к потомству, - изменения направлены, происходят закономерно, их можно предсказать, - имеют адаптивное (приспособительное) значение, - имеют массовый (групповой) и обратимый характер, - определенный фактор внешней среды вызывает сходное изменение у всех особей данного вида. Модификационная изменчивость имеет предел. Границы изменчивости признака, обусловленные генотипом называются нормой реакции. Она может быть узкой, когда признак изменяется незначительно (цвет глаз), и широкой, когда признак изменяется в широких пределах (рост, масса тела). В медицине часто приходится устанавливать норму реакции для оценки max и min количественных показателей (уровень гормонов, ферментов, гемоглобина и др.) Комбинативная изменчивость – это наследственная изменчивость, обусловленная перекомбинацией имеющихся генов и хромосом, без изменения структуры генов и хромосом (наследственного материала). Этот тип изменчивости проявляется уже на стадии образования половых клеток. Источниками комбинативной изменчивости являются процессы, происходящие в мейозе и в результате оплодотворения: 1. Рекомбинация генов при кроссинговере в профазе1 мейоза. 2. Рекомбинация хромосом в ходе мейоза (независимое расхождение хромосом и хроматид при мейозе) 3. Комбинация хромосом в результате слияния гамет при оплодотворении (случайное сочетание гамет при оплодотворении). Комбинативная изменчивость обеспечивает генотипическое разнообразие людей, объясняет наличие признаков у детей и внуков от родственников по отцовской и материнской линии. Мутационная изменчивость – способность генетического (наследственного) материала изменяться и эти изменения наследуются в потомстве. В основе мутационной изменчивости лежат мутации. Мутации – это внезапные изменения генетического материала под влиянием среды и передающиеся по наследству. Частота мутаций зависит от вида организма, от возраста, от фазы онтогенеза, стадии гаметогенеза, может происходить в половых и соматических клетках, иметь рецессивный и доминантный характер. Например, у человека до 6% гамет несут мутантные гены. Процесс образования мутаций называется мутагенезом. Факторы, вызывающие мутации называются мутагенными. Мутации первоначально действуют на генетический материал особи, а через генотип изменяется и фенотип. Мутагенные факторы Факторы, вызывающие мутации называются мутагенными факторами (мутагенами) и подразделяются на: 1. Физические; 2. Химические; 3. Биологические. К физическим мутагенным факторам относятся различные виды излучений, температура, влажность и др. Наиболее сильное мутагенное действие оказывает ионизирующее излучение – рентгеновские лучи, α-, β-, γ- лучи. Они обладают большой проникающей способностью. При действии их на организм они вызывают: а) ионизацию тканей – образование свободных радикалов (ОН) или (Н) из воды, находящейся в тканях. Эти ионы вступают в химическое взаимодействие с ДНК, расщепляют нуклеиновую кислоту и другие органические вещества; б) ультрафиолетовое излучение характеризуется меньшей энергией, проникает только через поверхностные слои кожи и не вызывает ионизацию тканей, но приводит к образованию димеров (химические связи между двумя пиримидиновыми основаниями одной цепочки, чаще Т-Т). Присутствие димеров в ДНК приводит к ошибкам при ее репликации, нарушает считывание генетической информации; в) разрыв нитей веретена деления; г) нарушение структуры генов и хромосом, т.е. образование генных и хромосомных мутаций. К химическим мутагенам относятся: - природные органические и неорганические вещества (нитриты, нитраты, алкалоиды, гормоны, ферменты и др.); - синтетические вещества, ранее не встречавшиеся в природе (пестициды, инсектициды, пищевые консерванты, лекарственные вещества). - продукты промышленной переработки природных соединений – угля, нефти. Механизмы их действия: а) дезаминирование – отщепление аминогруппы от молекулы аминокислот; б) подавление синтеза нуклеиновых кислот; в) замена азотистых оснований их аналогами. Химические мутагены вызывают преимущественно генные мутации и действуют в период репликации ДНК. К биологическим мутагенам относятся: - Вирусы (гриппа, краснухи, кори) - Невирусные паразитические организмы (грибы, бактерии, простейшие, гельминты) Механизмы их действия: а) вирусы встраивают свою ДНК в ДНК клеток хозяина. б) продукты жизнедеятельности паразитов-возбудителей болезней действуют как химические мутагены. Биологические мутагены вызывают генные и хромосомные мутации. Классификация мутаций Различают следующие основные типы мутаций: 1. По способу возникновения их подразделяют на спонтанные и индуцированные. Спонтанные – происходят под действием естественных мутагенных факторов внешней среды без вмешательства человека. Они возникают в условиях естественного радиоактивного фона Земли в виде космического излучения, радиоактивных элементов на поверхности земли. Индуцированные мутации вызываются искусственно воздействием определенных мутагенных факторов. 2. По мутировавшим клеткам мутации подразделяются на генеративные и соматические. Генеративные – происходят в половых клетках, передаются по наследству при половом размножении. Соматические – происходят в соматических клетках и передаются только тем клеткам, которые возникают из этой соматической клетки. Они не передаются по наследству. 3. По влиянию на организм: Отрицательные мутации – летальные (несовместимые с жизнью); полулетальные (снижающие жизнеспособность организма); нейтральные (не влияющие на процессы жизнедеятельности); положительные (повышающие жизнеспособность). Положительные мутации возникают редко, но имеют большое значение для прогрессивной эволюции. 4. По изменениям генетического материала мутации подразделяются на геномные, хромосомные и генные. Геномные мутации – это мутации, вызванные изменением числа хромосом. Могут появляться лишние гомологичные хромосомы. В хромосомном наборе на месте двух гомологичных хромосом оказываются три – это трисомия. В случае моносомии наблюдается утрата одной хромосомы из пары. При полиплоидии происходит кратное гаплоидному увеличение числа хромосом. Еще один вариант геномной мутации – гаплоидия, при которой остается только одна хромосома из каждой пары. Хромосомные мутации связаны с нарушением структуры хромосом. К таким мутациям относятся утраты участков хромосом (делеции), добавление участков (дупликация) и поворот участка хромосом на 180° (инверсия). Генные мутации, при которых изменения происходят на уровне отдельных генов, т.е. участков молекулы ДНК. Это может быть утрата нуклеотидов, замена одного основания на другое, перестановка нуклеотидов или добавление новых. |