Курс лекций по нормальной физиологии. Ю. И. Савченков. Красноярск Издво , 2012, 470 с
 Скачать 8.6 Mb. Скачать 8.6 Mb.
|

10.3. Функции новой коры, функциональное значение соматосенсорных и моторных зон коры больших полушарий.Кора больших полушарий головного мозга является наиболее молодым в филогенетическом отношении отделом мозга. Она представляет собой слой серого вещества, толщина которого колеблется от 1,5 до 3 мм. За счет большого количества складок площадь коры головного мозга составляет 1450-1700 кв.см. Кора больших полушарий является высшим интегративным центром регуляции процессов, протекающих в организме. Это доказывается характерными нарушениями, которые наблюдаются после декортикации. Такие животные уподобляются животным, находящимся на более низких стадиях эволюции. Причем чем более развито и совершенно животное, тем на большее количество ступеней эволюционного развития оно опускается. Максимальная деградация наблюдается у человека, лишенного коры (врожденно или в результате травмы), что, как правило, несовместимо с жизнью. Толщина новой коры колеблется от 1,5 до 4,5 мм и максимальна в передней центральной извилине. Число нейронов коры превышает 10 млрд. Первыми получили многие сведения о локализации функций в коре мозга морфологи и клиницисты. Морфологи показали, что цитоархитектоника коры изменяется не только по вертикали, но и по горизонтали. Это значит, что в разных участках коры она имеет специфическое строение. Морфологические особенности строения различных отделов коры мозга позволили разделить ее на несколько зон (карта Бродмана - 50 полей). Клиницисты доказали, что у человека многие участки коры больших полушарий имеют строго локализованные функции. Так, в области третьей левой лобной извилины расположен участок, относящийся в функции речи, в височной доле - центр слуха, в затылочной - зрения. Однако, в силу большой пластичности мозга и за счет перекрытия границ специфических зон в случае повреждения даже больших участков мозга функции этих отделов постепенно могут восстанавливаться. В настоящее время принято разделять кору на сенсорную, двигательную и ассоциативную. Сенсорные области. Корковые концы анализаторов имеютсвою топографию и на них проецируются определенные афференты проводящих систем. Корковые концы анализаторов разных сенсорных систем перекрываются. Помимо этого, в каждой сенсорной системе коры имеются полисенсорные нейроны, которые реагируют не только на «свой» адекватный стимул, но и на сигналы других сенсорных систем. Таламокортикальные пути проецируются на заднюю центральную извилину. Здесь имеется строгое соматотопическое деление. На верхние отделы этой извилины проецируются рецептивные поля кожи нижних конечностей, на средние — туловища, на нижние отделы — руки, головы. На заднюю центральную извилину в основном проецируются болевая и температурная чувствительность. В коре теменной доли (поля 5 и 7), где также оканчиваются проводящие пути чувствительности, осуществляется более сложный анализ: локализация раздражения, стереогноз. Зрительная система представлена в затылочной доле мозга: поля 17, 18, 19. Центральный зрительный путь заканчивается в поле 17; он информирует о наличии и интенсивности зрительного сигнала. В полях 18 и 19 анализируются цвет, форма, размеры, качества предметов. Поражение поля 19 коры большого мозга приводит к тому, что больной видит, но не узнает предмет (зрительная агнозия, при этом утрачивается также цветовая память). Слуховая система проецируется в поперечных височных извилинах (извилины Гешля), в глубине задних отделов латеральной (сильвиевой) борозды (поля 41, 42, 52). Именно здесь заканчиваются аксоны задних бугров четверохолмий и латеральных коленчатых тел. Обонятельная система проецируется в области переднего конца гиппокампальной извилины (поле 34). Кора этой области имеет не шести-, а трехслойное строение. При раздражении этой области отмечаются обонятельные галлюцинации, повреждение ее ведет к аносмии (потеря обоняния). Вкусовая система проецируется в гиппокампальной извилине по соседству с обонятельной областью коры (поле 43). Первичная соматосенсорная кора обеспечивает восприятие только простых ощущений, а анализ всей полноты сенсорного восприятия осуществляется многими отделами мозга в тесном взаимодействии с соматосенсорными зонами (при раздражении таких зон слышим звук, но не слово или музыкальную фразу). При разрушении первичных сенсорных зон возникает полная неспособность анализировать поступающую информацию (слепота, глухота и пр.). По всей границе с первичными сенсорными зонами (на расстоянии 1-5 см) располагаются зоны, называемые вторичными сенсорными зонами. Их разрушение вызывает заметное снижение способности мозга анализировать различные характеристики образов (потеря способности понимать значение слов, интерпретировать зрительные образы и т.п.). Особенно большое значение в этом плане имеет височная доля и угловая извилина. После обширного повреждения этих областей человек слышит и может различать слова, но не способен связать эти слова в законченную мысль, не может понимать их смысла. Электрическое раздражение этих областей у человека, находящегося в сознании, вызывает возникновение сложных мыслей, включая те, которые содержатся в его памяти. Отсюда следует, что у человека сложные образы, фиксированные в памяти, сохраняются в височной доле и угловой извилине. 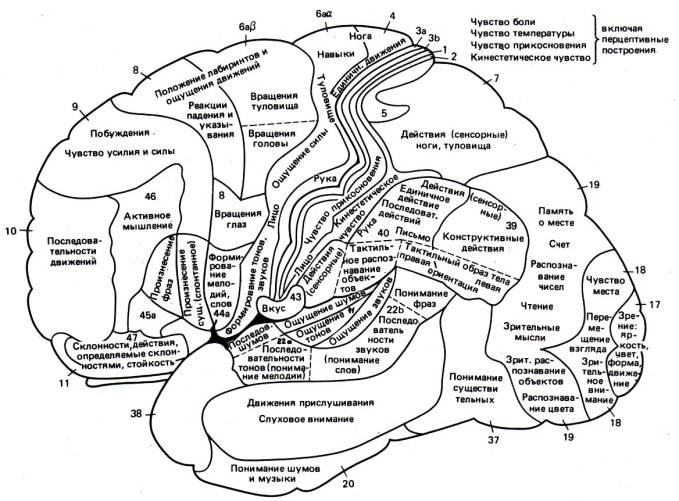 Рис. 14. Локализационная карта Клейста Моторные области. Впервые Фритч и Гитциг (1870) показали, что раздражение передней центральной извилины мозга (поле 4) вызывает двигательную реакцию. В то же время признано, что двигательная область является анализаторной. В передней центральной извилине зоны, раздражение которых вызывает движение, представлены по соматотопическому типу, но вверх ногами: в верхних отделах извилины — нижние конечности, в нижних — верхние. Спереди от передней центральной извилины лежат премоторные поля 6 и 8. Они организуют не изолированные, а комплексные, координированные, стереотипные движения. Эти поля также обеспечивают регуляцию тонуса гладкой мускулатуры, пластический тонус мышц через подкорковые структуры. В реализации моторных функций принимают участие также вторая лобная извилина, затылочная, верхнетеменная области. Двигательная область коры, как никакая другая, имеет большое количество связей с другими анализаторами, чем, видимо, и обусловлено наличие в ней значительного числа полисенсорных нейронов. Ассоциативные области. Все сенсорные проекционные зоны и моторная область коры занимают менее 20% поверхности коры большого мозга. Остальная кора составляет ассоциативную область. Каждая ассоциативная область коры связана мощными связями с несколькими проекционными областями. Считают, что в ассоциативных областях происходит ассоциация разносенсорной информации. В результате формируются сложные элементы сознания. Ассоциативные области мозга у человека наиболее выражены в лобной, теменной и височной долях. Каждая проекционная область коры окружена ассоциативными областями. Нейроны этих областей чаще полисенсорны, обладают большими способностями к обучению. Так, в ассоциативном зрительном поле 18 число нейронов, «обучающихся» условнорефлекторной реакции на сигнал, составляет более 60% от числа фоновоактивных нейронов. Для сравнения: таких нейронов в проекционном поле 17 всего 10—12%. Повреждение поля 18 приводит к зрительной агнозии. Больной видит, обходит предметы, но не может их назвать. В теменной ассоциативной области коры формируются субъективные представления об окружающем пространстве, о нашем теле. Это становится возможным благодаря сопоставлению соматосенсорной, проприоцептивной и зрительной информации. Лобные ассоциативные поля имеют связи с лимбическим отделом мозга и участвуют в организации программ действия при реализации сложных двигательных поведенческих актов. Первой и наиболее характерной чертой ассоциативных областей коры является мультисенсорность их нейронов, причем сюда поступает не первичная, а достаточно обработанная информация с выделением биологической значимости сигнала. Это позволяет формировать программу целенаправленного поведенческого акта. Вторая особенность ассоциативной области коры заключается в способности к пластическим перестройкам в зависимости от значимости поступающей сенсорной информации. Важной особенностью коры большого мозга является ее способность длительно сохранять следы возбуждения. Следовые процессы в спинном мозге после его раздражения сохраняются в течение секунды; в подкорково-стволовых отделах (в форме сложных двигательно-координаторных актов, доминантных установок, эмоциональных состояний) длятся часами; в коре мозга следовые процессы могут сохраняться по принципу обратной связи в течение всей жизни. Это свойство придает коре исключительное значение в механизмах ассоциативной переработки и хранения информации, накопления базы знаний. Сохранение следов возбуждения в коре проявляется в колебаниях уровня ее возбудимости; эти циклы длятся в двигательной области коры 3—5 мин, в зрительной — 5—8 мин. Основные процессы, происходящие в коре, реализуются двумя состояниями: возбуждением и торможением. Эти состояния всегда реципрокны. Они возникают, например, в пределах двигательного анализатора, что всегда наблюдается при движениях; они могут возникать и между разными анализаторами. Тормозное влияние одного анализатора на другие обеспечивает сосредоточенность внимания на одном процессе. Реципрокные отношения активности очень часто наблюдаются в активности соседних нейронов. Отношение между возбуждением и торможением в коре проявляется в форме так называемого латерального торможения. При латеральном торможении вокруг зоны возбуждения формируется зона заторможенных нейронов (одновременная индукция) и она по протяженности, как правило, в два раза больше зоны возбуждения. Латеральное торможение обеспечивает контрастность восприятия, что в свою очередь позволяет идентифицировать воспринимаемый объект. Помимо латерального пространственного торможения, в нейронах коры после возбуждения всегда возникает торможение активности и наоборот, после торможения — возбуждение — так называемая последовательная индукция. В тех случаях когда торможение не в состоянии сдерживать возбудительный процесс в определенной зоне, возникает иррадиация возбуждения по коре. Иррадиация может происходить от нейрона к нейрону, по системам ассоциативных волокон I слоя, при этом она имеет очень малую скорость — 0,5—2,0 м/с. В другом случае иррадиация возбуждения возможна за счет аксонных связей пирамидных клеток III слоя коры между соседними структурами, в том числе между разными анализаторами. Иррадиация возбуждения обеспечивает взаимоотношение состояний систем коры при организации условнорефлекторного и других форм поведения. Наряду с иррадиацией возбуждения, которое происходит за счет импульсной передачи активности, существует иррадиация состояния торможения по коре. Механизм иррадиации торможения заключается в переводе нейронов в тормозное состояние под влиянием импульсов, приходящих из возбужденных участков коры, например, из симметричных областей полушарий. 10.4. Вегетативная нервная система: особенности симпатического и парасимпатического отделов, их влияния. В обычных условиях внутренние органы в своей деятельности автономны, т.е. не подчиняются воле человека. По собственному желанию люди не могут изменить, например, работу печени, почек или желудка. Помимо того, что внутренние органы работают независимо от сознания, их деятельность протекает непрерывно, даже во время сна. В противоположность этому, деятельность поперечно-полосатой мускулатуры контролируется сознанием, т.е. является произвольной. Основываясь на этих различиях, французский анатом Биша еще в 17 веке подразделил нервную систему на два отдела. Ту ее часть, которая иннервирует внутренние органы, он предложил называть "вегетативной", а ту, которая иннервирует поперечно-полосатую мускулатуру - "анимальной". В конце 19 века Ленгли выделил в вегетативной нервной системе, которую он называл "автономной" две основные части - симпатический отдел и парасимпатический отдел. Вегетативная нервная система (ВНС) имеет свои, отличные от анимальной (соматической) нервной системы центры, афферентные и эфферентные проводники, хотя многие афферентные части являются общими. Характерным структурным отличием ВНС от соматической является наличие двух периферических нейронов - преганглионарного и постганглионарного, который представляет собой аналог мотонейрона, вынесенный на периферию, за пределы спинного мозга. В симпатической ВНС этой нейрон расположен в ганглии, в парасимпатической - интрамурально, в стенке иннервируемого органа. В последнее время кроме указанных двух отделов в составе ВНС стали выделять еще один - метасимпатическую, или энтериновую ВНС, представленную комплексом интрамуральных нервных образований в кишечнике, обеспечивающем сложную координацию гладкомышечных элементов кишечника при организации его моторики и перистальтики. В составе энтериновой (метасимпатической) НС имеются и чувствительные, и вставочные, и двигательные нейроны, между ними замыкаются многообразные короткие рефлекторные дуги. В строении и функциях вегетативной и симпатической НС имеется ряд характерных различий: 1. Наличие преганглионарного и постганглионарного нейрона в составе рефлекторной дуги ВНС. 2. Перерезка передних корешков спинного мозга вызывает различные изменения в эфферентной части соматической и вегетативной дуги. В соматической перерезка вызывает разобщение тела мотонейрона с его аксоном, что приводит к дегенерации последнего и развитию в тканях иннервируемого органа глубоких трофических нарушений и расстройства функции. На органы, иннервируемые ВНС, перерезка передних корешков оказывает меньшее влияние, так как дегенерации подвергается только преганглионарное волокно. Непосредственная иннервация тканей при этом не нарушается. Органы продолжают работать, трофика их не нарушается. Правда, при этом приходится учитывать, что механизмы, координирующие деятельность различных внутренних органов, выпадают, работа органа как бы децентрализуется, он начинает работать автономно, независимо от влияния вышележащих центров. Однако они продолжают получать импульсацию от вегетативных ганглиев и они поддерживают работу органов. Именно за это Ленгли и назвал ВНС автономной. 3.Следующее существенное отличие ВНС от СНС состоит в особенностях выхода волокон из мозга. Соматические волокна выходят из каждого сегмента ствола и спинного мозга. Вегетативные - сосредоточены избирательно в некоторых отделах - краниальном, торако-люмбальном и сакральном. Краниальный, в свою очередь, состоит из среднемозгового и бульбарного отделов. 4. Имеются отличия и в распределении вегетативных и соматических нервов на периферии. Соматические волокна имеют строгое сегментарное распределение, вегетативные нет. Большинство органов имеют двойную вегетативную иннервацию (симпатическую и парасимпатическую или энтериновую). 5. Волокна ВНС отличаются от волокон СНС меньшим диаметром и скоростью проведения возбуждения. Волокна ВНС имеют диаметр 8-5 микрон и скорость 1-3 м/сек. Соматические - соответственно 12-14 микрон и 70-120 м/сек. 6. Возбудимость соматических нервов выше, чем вегетативных: их хронаксия 0,1-0,8 мсек., тогда как у вегетативных 1,0-2,0 мсек. Вследствие более низкой возбудимости и меньшей скорости проведения возбуждения реакции ВНС имеют более медленный и инертный характер, чем реакции СНС. 7. Особую роль в функциях ВНС играют т.н. аксон рефлексы. и короткие вегетативные рефлексы. Аксон рефлексы отличаются от истинных тем, что при них не происходит передачи возбуждения с рецепторного нейрона на эффекторный. Они могут возникать в том случае, если аксоны пре- и постганглионарных нейронов ветвятся так, что одна ветвь иннервирует один орган, а другая - другой или другую часть органа. Аксон-рефлекторное происхождение имеет, например, расширение сосудов в области воспаления или при механическом (химическом) раздражении кожи (например, горчичником). Короткие вегетативные рефлексы замыкаются непосредственно между клетками Догеля 1 и 2 типа. 8. Характерным для ВНС является феномен широкой мультипликации. Суть этого явления состоит в том, что одно преганглионарное волокно способно образовывать синапсы на многих ганглионарных нейронах. При этом между числом пре- и постганглионарных волокон достигается соотношение 1:32. Центры ВНС. Низшими центрами, относящимися к центральному отделу ВНС, являются парасимпатические ядра кранио-бульбарного и сакрального отделов, представленные скоплениями клеточных тел эфферентных нейронов парасимпатической НС, аксоны которых выходят на периферию в составе 3,7,9, и 10 пар черепно мозговых и 1-4 крестцовых нервов. К низшим центрам ВНС относятся также симпатический центр Якубовича, расположенный в спинном мозгу от 1-2 грудного до 2-4 поясничных сегментов. Спинальные и краниальные вегетативные центры связаны с надсегментарными центрами. Так, в гипоталамусе имеются ядра, от которых зависит целый ряд вегетативных функций. Гипоталамус играет огромную роль в деятельности ВНС. Он является центром, осуществляющим интеграцию вегетативных процессов под регулирующим контролем корковых центров, с которыми он имеет обширные связи. Ядра гипоталамической области, которых в настоящее время насчитывают 32 пары, делятся на 3 группы: передние, средние и задние. Передняя группа имеет отношение к регуляции парасимпатической НС, а задняя - симпатической. Так, раздражение электрическим током гипоталамических ядер задней группы приводит к появлению признаков, характерных для возбуждения симпатического отдела ВНС - расширение зрачков, повышение активности пиломоторов, учащение сердцебиения, повышение АД, торможение двигательной активности ЖКТ и др. Напротив, раздражение гипоталамических ядер передней группы сопровождается появлением признаков, указывающих на преимущественное возбуждение парасимпатического отдела ВНС - ритм сердцебиений замедляется, время проведения по сердцу возбуждения возрастает, усиливается тонус и перистальтика ЖКТ, наступает сокращение мускулатуры мочевого пузыря и т.д. В условиях нормальной жизнедеятельности гипоталамус осуществляет регуляцию ВНС с помощью нисходящих эфферентных путей, начинающихся с этих групп ядер. Помимо прямых нервных влияний на деятельность внутренних органов, гипоталамус может оказывать на них воздействие и гуморальным путем. Последнее становится возможным благодаря нейросекреторным воздействиям гипоталамуса на гипофиз. В процессе нейросекреции в перинуклеарной зоне нейрона появляются мельчайшие гранулы, которые по мере накопления занимают все тело клетки, распространяясь и в ее отростки. Благодаря наличию в нейронах, связывающих супраоптическое ядро с гипофизом, этих гранул, этот тракт был назван нейросекреторным. В ножке гипофиза , помимо нервных волокон, имеются многочисленные кровеносные и лимфатические сосуды, по которым и осуществляется циркуляция нейросекретов. Если учесть, что помимо обширных связей с вышележащими центрами многие ядра гипоталамуса чувствительны к малейшим колебаниям внутренней среды, становится понятным исключительная роль гипоталамуса как образования, в котором осуществляется интеграция нервного и гуморального звена в едином процессе регуляции. Особое место среди нервных образований, принимающих участие в регуляции функций ВНС, принадлежит мозжечку. Мозжечок - высший надсегментарный орган, который регулирует и соматические и вегетативные функции (вегетативное обеспечение мышечных движений. Роль коры головного мозга в регуляции вегетативных функций доказывается не только возможностью произвольно изменять дыхание, но и наличием условно-рефлекторных реакций со стороны внутренних органов. Однако в обычных условиях в норме регуляция их деятельности осуществляется без участия коры мозга. Функции вегетативной нервной системы. И.П.Павловым было сформулировано понятие о тройственном характере функциональных влияний, которые может оказывать НС на ткани и органы: пусковое или корригирующее - приводит в деятельное состояние тот или иной орган или изменяет его функцию; сосудодвигательное - заключается в том, что путем изменения просвета сосудов увеличивается или уменьшается кровоснабжение данного органа; трофическое - за счет которого изменяется течение тканевого обмена веществ. В этом случае создаются благоприятные условия для работы органа. На фоне улучшенного функционального состояния пусковое нервное влияние оказывает больший эффект. Считают, что ВНС присущи все три вида влияний, с акцентом на второй и третий, тогда как СНС обладает преимущественно пусковой функцией. У ВНС первая функция более корригирующая, нежели пусковая. Одним из отличительных свойств ВНС является более активное выделение ею медиаторов и больший удельный вес их в передаче возбуждения на ткани. Все волокна ВНС подразделяются на адренэргические и холинэргические, выделяющие в своих окончаниях соответственно норадреналин или ацетилхолин. В последнее время найдены также т.н. пептидэргические волокна (медиатор пептидной природы). Показано, что парасимпатические волокна все являются холинэргическими. Симпатические преганглионары - холинэргические, постганглионары - адренэргические. Имеются и исключения. Так, периферические симпатические волокна, иннервирующие потовые железы, мозговое вещество надпочечников, а также сосуды сердца - холинэргические. Симпатические эффекты более длительны по времени и захватывают обширные области. Парасимпатические - более кратковременны и более локальны, т.к. холинэстераза более активна, чем моноаминоксидаза, разрушающая норардреналин. Универсальный характер симпатической иннервации. Симпатическая система иннервирует все внутренние органы. Кроме того, она иннервирует разные отделы ЦНС: волокна ее отделяются от превертебральных ганглиев и через задние корешки направляются в спинной мозг, проходя там вверх и вниз. Попадают они и в головной мозг, иннервируя его отделы. Получают симпатическую иннервацию и поперечно-полосатые мышцы. Находящиеся в коже образования - мышечные волокна, приводящие в движение кожные волосы (пиломоторы), потовые железы - все получают симпатические веточки. Все рецепторы, воспринимающие то или иное раздражение, также снабжены симпатическими ответвлениями. Кроме того, кровеносные сосуды повсеместно имеют симпатическую иннервацию, и , следовательно, вегетативные нервы проникают во все органы и вместе с сосудами, особенно это относится к симпатическим волокнам, которые распространяются по периартериальным нервным сплетениям. Каков же смысл столь обильной симпатической иннервации? Адаптационно-трофическое влияние НС. Об участии нервной системы в трофике, т.е. в питании тканей, известно со времен И.П.Павлова, который обнаружил, что один из нервов сердечного сплетения, не изменяя ритма работы сердца, увеличивает интенсивность сердечных сокращений. В лаборатории Орбели были проведены классические опыты по изучению влияния симпатической нервной системы на мышечную ткань. Оказалось, что если раздражением двигательного нерва довести мышцу до утомления, а затем раздражать подходящие к ней симпатические нервы, то присоединение подобного раздражения приводит к активации сокращений утомившейся мышцы (феномен Орбели-Гинецинского). Было доказано специальными опытами, что такое повышение работоспособности мышцы не связано с улучшением кровообращения, а является результатом стимуляции обменных процессов. Местом приложения этого симпатического влияния является мионевральная пластинка. Симпатическая нервная система оказывает влияние на функции ЦНС во многих ее звеньях. Оказалось, что раздражение симпатического нерва влияет на скорость рефлекторных реакций спинного мозга. Удаление шейных симпатических узлов у собаки вызывает заметные нарушения ее ВНД. Симпатические волокна могут значительно изменять и чувствительность рецепторов, т.е. эффекты ее схожи с деятельностью РФ ствола мозга. Автономность ВНС. Ленгли недаром назвал вегетативную нервную систему автономной. Опыты показали, что если у животного полностью убрать спинной мозг, то при этом, несмотря на полную неподвижность, вегетативные функции - пищеварение, работа желез, почек и др. продолжают осуществляться, и при хорошем уходе такие животные могут жить довольно долго. Следовательно, ганглии ВНС в какой-то мере самостоятельно могут обслуживать нужны организма. Эта самостоятельность находит проявление и в виде т.н. местных рефлекторных дуг, функционирующих во внутренних органах независимо от ЦНС (сердце, ЖКТ). Таким местным рефлекторным дугам приписывают, например, напряжение стенки кишечника при попадании в его полость комка пищи. Подобным же образом возникает и секреторный ответ. Это - работа энтериновой нервной системы кишечника, расположенной в Ауэрбаховском и Мейснеровском нервных сплетениях. Значительную роль в деятельности внутренних органов играют висцеральные и аксон рефлексы. Эти рефлексы частью могут быть висцеро-моторные и висцеросенсорные. Примером висцеро-моторного рефлекса является напряжение прямой мышцы живота при поражении внутренних органов, раздражении брюшины, например, при полостном кровотечении. Чувствительные окончания в брюшине посылают импульсы, переходящие на ветви симпатических нервов, вступающие в связь с мускулатурой брюшной стенки, которая напрягается и длительно остается в таком положении. Примером висцеросенсорного рефлекса являются отраженные боли в коже при поражении определенных внутренних органов. Взаимоотношения между симпатической и парасимпатической НС. В ряде случаев влияния, оказываемые симпатической и парасимпатической НС носят противоположный характер. Однако, следует помнить, что такие "антагонистические" отношения проявляются не всегда и не везде. В ряде органов функциональный антагонизм отсутствует. Так, нельзя говорить об антагонистических отношениях симпатических нервов, расширяющих зрачок, и парасимпатических, суживающих его. В этом случае оба типа волокон оказывают стимулирующее влияние, но на разные мышцы. Даже тогда, когда орган имеет и симпатическую, и парасимпатическую иннервацию, антагонизм часто отсутствует. Так, для слюнных желез секреторным для жидкой фазы является парасимпатикус, а для ферментов - симпатикус. |