Курс лекций по нормальной физиологии. Ю. И. Савченков. Красноярск Издво , 2012, 470 с
 Скачать 8.6 Mb. Скачать 8.6 Mb.
|

|
Моторная функция желудка. Во время и в первые минуты после приема пищи желудок расслабляется — наступает пищевая рецептивная релаксация желудка, которая способствует депонированию пищи в желудке и его секреции. Спустя некоторое время в зависимости от вида пищи сокращения усиливаются, при этом наименьшая сила сокращения отмечается в кардиальной части желудка и наибольшая — в антральной. Сокращения желудка начинаются на большой кривизне в непосредственной близости от пищевода, где находится кардиальный водитель ритма. Второй водитель ритма локализован в пилорической части желудка. При баллонной гастрографии регистрируется три типа волн сокращений желудка: I — однофазные волны низкой амплитуды, давление колеблется от 1—2 до 5—10 мм рт. ст., длительностью 5—20 с; II — однофазные волны большой амплитуды, давление составляет 40—80 мм рт. ст., длительностью 12— 60 с; III — сложные волны, возникают на фоне меняющегося исходного давления. Волны I и II типов носят перистальтический характер, поддерживают определенный тонус желудка, обеспечивают смешивание пищи с желудочным соком в непосредственной близости к слизистой оболочке желудка. Средняя частота этих волн 3 в 1 мин. В центральной части желудка содержимое не перемешивается, поэтому пища, принятая в разное время, располагается в желудке слоями (стратификация). Волны III типа характерны для пилорической части желудка, носят пропульсивный характер и участвуют в эвакуации содержимого в двенадцатиперстную кишку. 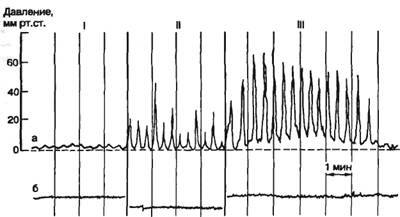 Рис. 47. Три типа волн сокращений желудка здорового человека (I—III). а — сокращения желудка; б — пневмограмма. При регистрации внутрижелудочного давления методом открытых катетеров выявляются сокращения желудка двух типов: фазовые (тип А) и тонические (тип Б). Волны типа А быстрые, перистальтические, продолжительностью 10—20 сек с частотой около 3 в 1 мин, волны типа Б — медленные, тонические, длительностью до 2 мин. Волны типа А бывают двух видов, первые имеют амплитуду и колебание давления от 1 до 15 мм рт. ст., у вторых амплитуда и колебание давления 16—30 мм рт. ст. Тонические волны (тип Б) могут сочетаться и не сочетаться с фазовыми. В наполненном пищей желудке возникают три основных вида движений: перистальтические волны, систолические сокращения пилорического отдела и тонические, уменьшающие размер полости дна и тела желудка. Частота перистальтических сокращений около 3 в 1 мин; они распространяются от кардиальной части желудка к пилорической со скоростью около 1 см/с, быстрее по большой, чем по малой кривизне, длятся около 11/2 с. В пилорической части скорость распространения перистальтической волны увеличивается до 3—4 см/с. После приема пищи и в зависимости от ее вида параметры моторной деятельности желудка приобретают характерную динамику. В течение первого часа перистальтические волны слабые, в дальнейшем они усиливаются (в пилорическом отделе увеличиваются их амплитуда и скорость распространения), проталкивая пищу к выходу из желудка. Давление в пилорическом отделе повышается до 10—25 см вод. ст., открывается сфинктер привратника (пилорический сфинктер), и порция желудочного содержимого переходит в двенадцатиперстную кишку. Оставшееся (большее) количество его возвращается в проксимальную часть пилорического отдела желудка. Такие движения желудка обеспечивают перемешивание и перетирание (фрикционный эффект) пищевого содержимого, его гомогенизацию. Характер, интенсивность, временная динамика моторики зависят от количества и вида пищи, от эффективности ее переваривания в желудке и кишечнике, обеспечивается регуляторными механизмами. Перистальтическим называют циркулярное сокращение полосы мышц желудка, движущееся в каудальном направлении. Ширина продвигающейся полосы сокращения - 1,5-2 см. Возникает оно в области кардии и продвигается до нижней трети желудка. Смещаются только поверхностные слои содержимого. В кардии есть пейсмекер ритма. Распространяется возбуждение по косому слою. Нервы принимают участие в координации этих сокращений. Частота 3-4 /мин. и не зависит ни от каких условий. Сила - небольшая ("рябь на воде"). Скорость распространения может изменяться и зависит от состояния желудка. В теле она равна около 1 см/сек, в пилорическом отделе 3-4 см/сек. При пилоростенозе - 6-1 см/сек, при гастрите - 70-80 см/сек. Несмотря на значительные отличия в длине большой и малой кривизны (в 3 раза), перистальтическая волна достигает пилоруса одновременно по периметру всего желудка. Как только волна перемещается на терминали антрального сегмента, этот сегмент сильно сокращается как функциональная единица. Такой тип сокращения называется антральная систола. В время этого сокращения значительная часть содержимого антрум переходит в 12-перстную кишку. Тонические волны - это передвигающие сокращения. Они оказывают давление на значительно большее количество содержимого желудка, чем перистальтические. Уплотняют и сдвигают содержимое фундального отдела в дистальную часть, а также имеют значение для более эффективной обработки и эвакуации. Тоническое сокращение - это не передвигающее повышение тонуса мышц желудка, которое приводит к уменьшению полости и к повышению давления в желудке. Длительность - до нескольких минут. Способствуют перемешиванию содержимого и пропитыванию его соком. Регуляция моторики желудка. Раздражение блуждающих нервов и выделение АХ усиливают моторику желудка: увеличивают ритм и силу сокращений, ускоряют движение перистальтических волн. Влияния блуждающих нервов могут оказывать и тормозной эффект: рецептивная релаксация желудка, снижение тонуса пилорического сфинктера. Раздражение симпатических нервов и активация α-адренорецепторов тормозят моторику желудка: уменьшают ритм и силу его сокращений, скорость движения перистальтической волны. В регуляции моторики желудка велико значение гастроинтестинальных гормонов. Моторику желудка усиливают гастрин, мотилин, серотонин, инсулин, а тормозят — секретин, ХЦК, глюкагон, ЖИП, ВИП. Механизм их влияний на моторику прямой (непосредственно на мышечные пучки и миоциты) и опосредованный через интрамуральные нейроны. Моторика желудка зависит от уровня его кровоснабжения и сама влияет на него, изменяя сопротивление кровотоку при сокращениях желудка. Эвакуация содержимого желудка в 12-перстную кишку. Скорость эвакуации пищи из желудка зависит от многих факторов: объема, состава и консистенции, величины осмотического давления, температуры и рН содержимого желудка, градиента давления между полостями пилорического отдела желудка и двенадцатиперстной кишки, состояния сфинктера привратника, аппетита, с которым принималась пища, состояния водно-солевого гомеостаза и ряда других причин. Пища, богатая углеводами, при прочих равных условиях быстрее эвакуируется из желудка, чем богатая белками. Жирная пища эвакуируется из него с наименьшей скоростью. Жидкости начинают переходить в кишку сразу после их поступления в желудок. Время полной эвакуации смешанной пищи из желудка здорового взрослого человека составляет 6—10 ч. Регуляция скорости эвакуации содержимого желудка осуществляется рефлекторно при активации рецепторов желудка и двенадцатиперстной кишки. Раздражение механорецепторов желудка ускоряет эвакуацию его содержимого, а двенадцатиперстной кишки — замедляет. Из химических агентов, действующих на слизистую оболочку двенадцатиперстной кишки, значительно замедляют эвакуацию кислые (рН меньше 5,5) и гипертонические растворы, 10 % раствор этанола, глюкоза и продукты гидролиза жира. Скорость эвакуации зависит также от эффективности гидролиза питательных веществ в желудке и тонкой кишке; недостаточность гидролиза замедляет эвакуацию. Следовательно, желудочная эвакуация «обслуживает» гидролитический процесс в двенадцатиперстной и тонкой кишке и в зависимости от хода его с различной скоростью «загружает» основной «химический реактор» пищеварительного тракта — тонкую кишку. Регуляторные влияния на моторную функцию гастродуоденального комплекса передаются с интеро- и экстероцепторов через ЦНС и короткие рефлекторные дуги, замыкающиеся в экстра- и интрамуральных ганглиях. В регуляции эвакуаторного процесса принимают участие гастроинтестинальные гормоны, влияющие на моторику желудка и кишечника, изменяющие секрецию главных пищеварительных желез и через нее — параметры эвакуируемого желудочного содержимого и кишечного химуса. Методы исследования: у человека - однократное или фракционное зондирование, рентгенография, УЗИ, электрогастрография. 26. 2. Пищеварение в 12-ти перстной кишке: общая характеристика, рефлекторная и гормональная функция. Роль поджелудочной железы в пищеварении (состав, количество, ферменты сока поджелудочной железы), регуляция панкреатической секреции. Пищеварение в 12-перстной кишке. В обеспечении начального этапа пищеварения большая роль принадлежит процессам, происходящим в двенадцатиперстной кишке. Натощак ее содержимое имеет слабощелочную реакцию (рН 7,2—8,0). При переходе в кишку порций кислого содержимого желудка реакция содержимого двенадцатиперстной кишки становится кислой, но затем она сдвигается к нейтральной за счет поступающих в кишку щелочных секретов поджелудочной железы, тонкой кишки и желчи, которые прекращают действие желудочного пепсина. В инактивации пепсина велика роль желчи. У человека рН содержимого двенадцатиперстной кишки колеблется в пределах 4—8,5. Чем выше его кислотность, тем больше выделяется сока поджелудочной железы, желчи и кишечного секрета, замедляется эвакуация содержимого желудка в двенадцатиперстную кишку и ее содержимого в тощую кишку. По мере продвижения по двенадцатиперстной кишке пищевое содержимое смешивается с поступающими в кишку секретами. Химус находится в 12-п. кишке очень короткое время, поэтому о какой-либо переработке ее в полости 12-п.кишки говорить нельзя. Поступающая в дуоденум пищевая масса (химус) подвергается действию поджелудочного сока, желчи, а также сока бруннеровых и либеркюновых желез 12-п.кишки и желчи. Вне пищеварения содержимое 12-п.кишки имеет слабо - щелочную реакцию (рН 7,2-8,0). При переходе в нее порций кислого желудочного содержимого реакция в кишке становится кислой, а затем постепенно нормализуется. У человека в связи с этим реакция в кишке колеблется от 4.0 до 8,5 рН. Несмотря на то, что какой-либо реальной химической переработки пищевой массы в 12-п. кишке не происходит, роль ее в пищеварительном процессе не ограничивается тем, что в ее полости происходит смешивание химуса с соками поджелудочной железы и желчью. 12-перстная кишка является важнейшей рефлексогенной зоной, с рецепторов которой начинаются рефлексы, обеспечивающие порционную эвакуацию содержимого желудка, стимуляцию. выделения желчи и поджелудочного сока, стимуляцию моторики тонкой кишки. Наконец, очень важной функцией 12-перстной кишки является гормональная. В клетках ее слизистой вырабатываются гормоны и биологически активные вещества, осуществляющие регуляцию секреторной и моторной деятельности всех других отделов ЖКТ. Перечень некоторых этих веществ и их эффекты представлены в таблицах 1 и 2. Дело в том, что в управлении пищеварительными функциями принимают участие пептиды и амины, которые продуцируются эндокринными клетками самого пищеварительного тракта. Эти клетки рассеяны в слизистой оболочке и пищеварительных железах и в совокупности составляют диффузную эндокринную систему. Особенно много таких клеток в 12-перстной кишке. Продукты их деятельности называют гастроинтестинальными гормонами, энтеринами, регуляторными пептидами пищеварительного тракта. Это не только пептиды, но и амины. Некоторые из них продуцируются и нервными клетками ЖКТ. В первом случае эти биологически активные вещества выступают в роли гормонов (доставляются к органам-мишеням общим и региональным кровотоком) и парагормонов (диффундируют через интерстициальную ткань к рядом или недалеко расположенной клетке). Во втором случае эти вещества играют роль нейротрансмиттеров. Открыто более 30 регуляторных пептидов пищеварительного тракта, некоторые из них существуют в нескольких изоформах, отличаясь числом аминогрупп и физиологической активностью. Гастроинтестинальные гормоны имеют широкий спектр физиологической активности, влияя на пищеварительные функции и вызывая общие эффекты. В пищеварительном тракте пептиды и амины стимулируют, тормозят, модулируют секрецию, моторику, всасывание, оказывают трофические влияния, в том числе воздействуя на пролиферативные процессы, например изменяют количество гландулоцитов в слизистой оболочке желудка и поджелудочной железе, уменьшая или увеличивая их массу. Каждый из регуляторных пептидов вызывает несколько эффектов, один из которых часто является основным. Ряд пептидов выступает в роли рилизинг-факторов для других пептидов, которые вызывают изменения пищеварительных функций в таком регуляторном каскаде. Эффекты регуляторных пептидов зависят от их дозы, механизмов, с помощью которых была стимулирована функция. Регуляторные пептиды относятся к числу «короткоживущих» веществ (период полураспада несколько минут), вызываемые ими эффекты, как правило, значительно длительнее. Пищеварительная функция поджелудочной железы. Основную массу поджелудочной железы (80—85 %) составляют экзокринные элементы, среди которых 80—95 % приходится на ацинарные клетки; эти клетки секретируют ферменты (и небольшое количество неферментных белков); центроацинозные и протоковые клетки секретируют воду, электролиты, слизь; из протоков компоненты смешанного секрета частично реабсорбируются. Поджелудочная железа человека натощак выделяет небольшое количество секрета. При поступлении пищевого содержимого из желудка в двенадцатиперстную кишку поджелудочная железа человека выделяет сок со средней скоростью 4,7 мл/мин. За сутки выделяется 1,5—2,5 л сока сложного состава. Сок представляет собой бесцветную прозрачную жидкость со средним содержанием воды 987 г/л. Щелочная среда сока (рН 7,5—8,8) обусловлена наличием в нем гидрокарбонатов (до 150 ммоль/л). Концентрация гидрокарбонатов в соке изменяется прямо пропорционально скорости секреции. В соке содержатся хлориды натрия и калия; между концентрацией гидрокарбонатов и хлоридов обратная зависимость. Гидрокарбонаты сока поджелудочной железы участвуют в нейтрализации и ощелачивании кислого пищевого содержимого желудка в двенадцатиперстной кишке. В соке отмечается значительная концентрация белка, основную часть которого составляют ферменты. Сок поджелудочной железы богат ферментами, которые синтезируются в ацинозных панкреоцитах. К ферментам поджелудочной железы относятся α-амилаза, прокарбоксипептидазы, трипсиноген, химотрипсиноген, проэластаэы, колипазы, профосфолипаза, нуклеазы. Ферменты поджелудочного сока переваривают все виды питательных веществ. Амилаза, липаза и нуклеаза секретируются поджелудочной железой в активном состоянии, а протеазы — в виде зимогенов. Основные эффекты влияния гастроинтестинальных гормонов на пищеварительные функции.
Трипсиноген сока поджелудочной железы в двенадцатиперстной кишке под действием ее фермента энтерокиназы превращается в трипсин. Последующую активацию трипсиногена вызывает трипсин. Активация состоит в отщеплении от трипсиногена гексапептида под действием энтерокиназы и трипсина при рН 6,8—8,0. Процесс ускоряется в присутствии ионов Са2+. Химотрипсиноген активируется трипсином. Трипсин и химотрипсин (а также панкреатопептидаза, или эластаза) расщепляют преимущественно внутренние пептидные связи белков. Эти ферменты действуют и на высокомолекулярные полипептиды, в результате чего образуются низкомолекулярные пептиды и аминокислоты. В составе сока поджелудочной железы выделяется некоторое количество ингибитора трипсина. Поджелудочная железа синтезирует прокарбоксипептидазы А и В, проэластазы и профосфолипазу. Они активируются трипсином с образованием соответствующих ферментов: карбоксипептидаз А и В, эластаз и фосфолипазы. Сок поджелудочной железы богат α-амилазой, расщепляющей полисахариды до ди- и моносахаридов. На производные нуклеиновых кислот действуют рибо- и дезоксирибонуклеазы. Панкреатическая липаза расщепляет жиры, в основном триглицериды, до моноглицеридов и жирных кислот. На липиды действуют также фосфолипаза А2 и эстераза. Поджелудочная железа секретирует профермент — панкреатическую фосфолипазу, которая активируется трипсином. Поскольку триглицериды нерастворимы в воде, липаза действует только на поверхности жира. Чем больше суммарная площадь поверхности контакта жира и липазы, тем активнее идет его гидролиз. Поэтому эмульгирование жира имеет огромное значение для его переваривания. Эмульгирование обеспечивается желчью, точнее — ее желчными кислотами и их солями. Размеры частиц жира 0,2—5,0 мкм. Активность липазы повышает также фермент колипаза. Она связывается с липазой в присутствии желчных солей и снижает оптимум рН действия фермента с 9 до 6—7, а также способствует адсорбции липазы на слизистой оболочке кишки. Повышают активность липазы присутствие и ионов Са2+. Под действием липаз осуществляется обычно неполный гидролиз триглицеридов; при этом образуется смесь из моноглицеридов (около 50 %), жирных кислот и глицерина (40 %), ди- и триглицеридов (3—10%). Методы исследования секреции поджелудочной железы - выведение протока наружу по Павлову или по Орлову (лучше). У человека получить чистый сок трудно, при зондировании получают смесь соков. При эндоскопическом исследовании возможно проникнуть в проток, но это возможно не всегда. Секреция поджелудочного сока начинается через 2-3 минуты после приема пищи и продолжается 6-14 часов в зависимости от состава пищи. Натощак поджелудочный сок выделяется только в небольших количествах во время периодической деятельности пищеварительного тракта. Количество сока и его ферментный состав зависят от качества поступающего химуса. Наибольшее количество сока выделяется на мясо на втором часу, на хлеб - на первом часу, на молоко - на третьем часу, т.е. так же, как и желудочный сок. При мясной пище, содержащей мало жира, у человека выделятся в 2,5 раза больше сока, чем при богатой жирами пище. При изменении характера питания меняется и ферментный состав сока. Регуляция секреции поджелудочной железы. Секреция поджелудочной железы регулируется нервными и гуморальными механизмами. Нервная регуляция. Раздражение блуждающего нерва вызывает выделение большого количества сока поджелудочной железы, богатого ферментами. Холинергические волокна блуждающих нервов с помощью АХ действуют на М-холинорецепторы панкреацитов. Затем высвобождающиеся ион Са2+ и комплекс ГЦ — цГМФ в роли вторичных мессенджеров стимулируют секрецию панкреоцитами ферментов и гидрокарбонатов. Холинергические нейроны, кроме того, потенцируют секреторные эффекты секретина и ХЦК. Хирургическая ваготомия существенно снижает секрецию поджелудочной железы. Симпатические волокна, иннервирующие поджелудочную железу через β-адренорецепторы, тормозят ее секрецию, усиливают синтез органических веществ в ней. Адренергические эффекты снижения секреции обеспечиваются также уменьшением кровоснабжения поджелудочной железы путем сужения кровеносных сосудов через их α-адренорецепторы. Торможение секреции вызывают болевые раздражения, сон, напряженная физическая и умственная работа и др. Гуморальная регуляция. Первым открытым (и названным "гормоном") явился секретин — стимулятор обильного сокоотделения и секреции гидрокарбонатов. Высвобождение этого гормона в кровь S-клетками двенадцатиперстной кишки происходит при действии на ее слизистую оболочку перешедшего в кишку кислого желудочного содержимого. Секретин через соответствующие мембранные рецепторы и вторичные мессенджеры АЦ — цАМФ в большей мере стимулирует секрецию центроацинозных и протоковых клеток, в меньшей мере — ацинозных клеток, поэтому выделяется секрет с высокой концентрацией гидрокарбонатов и низкой ферментативной активностью. Вторым гормоном, усиливающим секрецию поджелудочной железы, является холецистокинин (ХЦК). Высвобождение гормона в кровь из клеток слизистой оболочки двенадцатиперстной и тощей кишки происходит под влиянием пищевого химуса (особенно продуктов начального гидролиза пищевых белков и жиров, углеводов, некоторых аминокислот). Стимулируют высвобождение ХЦК присутствие ионов Са2+ и снижение рН в 12-перстной кишке. ХЦК действует преимущественно на ацинусы поджелудочной железы, поэтому выделяющийся в ответ на стимуляцию этим гормоном сок богат ферментами. Одновременное действие на железу секретина и ХЦК (при приеме пищи) усиливает их стимуляторный эффект. Секретин и ХЦК применяются в клинике как стимуляторы секреции при диагностике заболеваний поджелудочной железы. Пептид химоденин стимулирует секрецию химотрипсиногена. Секреция поджелудочной железы усиливается также гастрином, серотонином, инсулином, бомбезином, солями желчных кислот. Тормозят выделение поджелудочного сока глюкагон, соматостатин, вазопрессин, вещество Р, АКТГ, энкефалин, кальцитонин, ЖИП, ПП, УУ. ВИП может возбуждать и тормозить секрецию поджелудочной железы. Фазы секреции поджелудочной железы. Секреция сока поджелудочной железы резко усиливается через 2—3 мин после приема пищи и продолжается 6—14 ч. От количества и качества пищи зависят объем, состав выделяющегося сока, динамика выделения. Чем выше кислотность пищевого содержимого желудка, поступающего в двенадцатиперстную кишку, тем больше выделяется сока поджелудочной железы и тем больше гидрокарбонатов в его составе. Поэтому кривая секреции поджелудочной железы в некоторой мере повторяет кривую желудочного сокоотделения. Отличия кривых секреции желудка и поджелудочной железы определяются в основном буферными свойствами пищи, которая частично нейтрализует кислоту желудочного сока, и скоростью эвакуации содержимого желудка в двенадцатиперстную кишку. Фазы секреции поджелудочной железы при стимуляции ее приемом пищи те же, что и при регуляции желудочной секреции, но в отличие от них более выражены гормональные влияния на поджелудочную железу, особенно в кишечную фазу. Секреция имеет характерную динамику, зависящую от вида принятой пищи. Первая, или мозговая, фаза секреции поджелудочной железы обусловлена видом, запахом пищи и другими раздражителями, связанными с приемом пищи (условно-рефлекторные раздражения), а также воздействиями на рецепторы слизистой оболочки рта, жеванием и глотанием (безусловно-рефлекторные раздражения). Нервные импульсы, возникающие в рецепторах, достигают продолговатого мозга и затем по волокнам блуждающего нерва поступают к железе и вызывают ее секрецию. У человека с фистулой протока поджелудочной железы наблюдали условно-рефлекторное выделение сока поджелудочной железы через 2—3 мин после того, как испытуемому говорили о пище, которую ему дадут. В реализации первой фазы секреции принимают участие и регуляторные пептиды, высвобождение которых стимулировалось рефлекторными механизмами блуждающих нервов. Вторая, или желудочная, фаза характеризуется тем, что секреция во время нее стимулируется и поддерживается путем ваговагального рефлекса с механо- и хеморецепторов желудка и с помощью гастрина. С переходом желудочного содержимого в двенадцатиперстную кишку начинается третья, или кишечная, фаза секреции поджелудочной железы. В эту фазу секреция стимулируется ваговагальным дуоденопанкреатическим рефлексом, но ведущее значение имеет высвобождение в кровь секретина и ХЦК. Высвобождение их происходит при действии на слизистую оболочку 12перстной кишки кислого ее содержимого. Чем больше свободных ионов Н+, тем больше высвобождается секретина и тем выше объем сока поджелудочной железы и секреция гидрокарбонатов. Гидрокарбонаты связывают ионы Н+, что приводит к повышению рН среды и уменьшает высвобождение секретина и соответственно объем сока и секрецию гидрокарбонатов. Секреция ферментов в кишечную фазу стимулируется ХЦК и рефлекторно за счет ваговагального рефлекса. В кишечную фазу велика роль саморегуляции секреции поджелудочной железы по принципу отрицательной обратной связи в зависимости от свойств содержимого двенадцатиперстной кишки. Установлено, что отведение сока поджелудочной железы из двенадцатиперстной кишки вызывает гиперсекрецию поджелудочной железы, обратное введение сока в кишку тормозит эту секрецию. Введение в кишку гидрокарбонатов снижает объем секреции, концентрацию и дебит (выделение) гидрокарбонатов в составе сока. Таким образом, свойства секрета поджелудочной железы в кишечную фазу в большой мере определяются соотношением в химусе двенадцатиперстной кишки ферментов и гидролизуемых ими субстратов: избыток ферментов селективно тормозит их секрецию, избыток субстрата снимает эти тормозные влияния, и продукты гидролиза субстрата стимулируют секрецию соответствующих ферментов поджелудочной железой. Этот механизм направлен на срочную адаптацию секреции ферментов поджелудочной железы к виду принятой пищи. Его реализация обеспечивается М-холинергическими и (β-адренергическими влияниями, ХЦК, секретином. В целом нервные влияния при приеме пищи обеспечивают пусковые воздействия на поджелудочную железу, в последующей коррекции ее секреции большую роль играют гуморальные механизмы. Однако высвобождение гормонов 12-перстной кишкой и действие их на поджелудочную железу более выражены при сохраненной ее иннервации, что подчеркивает единство нервных и гуморальных механизмов регуляции секреции поджелудочной железы. При стимуляции секреции поджелудочной железы усиливается ее кровоснабжение, что важно для поддержания секреции на высоком уровне. Секреция гуморально корригируется и всосавшимися питательными веществами. Эти влияния осуществляются непосредственно на панкреоциты, выражено влияние на них некоторых аминокислот и глюкозы, опосредуются через центральные нервные механизмы (например, гипоталамический и бульварный центры автономной нервной системы) и регуляторные пептиды. Влияние пищевых режимов на секрецию поджелудочной железы. Прием пищи вызывает увеличение выделения всех ферментов в составе сока, но для разных видов пищи это увеличение выражено в разной мере. В случае приема пищи с большим содержанием углеводов в наибольшей мере увеличивается секреция амилазы, белков — трипсина и химотрипсина, прием жирной пищи вызывает секрецию сока с более высокой липолитической активностью. Спектр ферментов сока поджелудочной железы срочно адаптируется к виду принимаемой пищи во все три фазы секреции и особенно в кишечную фазу. Существуют и медленные адаптации секреции ферментов в зависимости от длительного режима питания. Суть адаптации состоит в том, что поджелудочная железа синтезирует и секретирует больше того фермента, который гидролизует преобладающие в рационе питательные вещества. Преобладание в рационе белков повышает выделение в составе сока поджелудочной железы протеаз, преимущественное углеводное питание вызывает увеличение выделения с соком амилазы, а большое количество жира в рационе — выделение с соком липазы. Эти изменения носят согласованный с изменениями функционирования других отделов пищеварительного тракта характер, составляя часть интегрированных адаптаций всего пищеварительного конвейера. |