Курс лекций по нормальной физиологии. Ю. И. Савченков. Красноярск Издво , 2012, 470 с
 Скачать 8.6 Mb. Скачать 8.6 Mb.
|

ЛЕКЦИЯ 31. ГОМЕОСТАТИЧЕСКИЕ ФУНКЦИИ ПОЧЕК.31-1. Концентрация мочи в почках. Поворотно-противоточная система почки. Значение дистальных извитых канальцев и собирательных трубок нефрона в формировании конечной мочи. гормональный механизм регуляции этих процессов.Способностью к образованию мочи с большей осмотической концентрацией, чем кровь, обладают лишь почки теплокровных животных. Образование осмотически концентрированной мочи обусловлено деятельностью поворотно-противоточной множительной системы в почке, которая представлена параллельно идущими коленами петли Генле и собирательными трубочками. Моча двигается в этих канальцах в противоположных направлениях, а процессы транспорта веществ в одном колене системы усиливаются – «умножаются» – за счет деятельности другого колена. Важнейшую роль в противоточном механизме играет активный перенос Na+ и Cl– из просвета восходящего колена петли Генле в межклеточную жидкость. Принципиально, что восходящее колено по всей его длине непроницаемо для воды. Вследствие выхода NaCl жидкость в просвете восходящего колена становится гипотоничной, а межклеточная жидкость – гипертоничной. Принцип противоточного обмена достаточно широко распространен в природе и используется в технике. Механизм работы такой системы рассмотрим на примере кровеносных сосудов в конечностях арктических животных. Во избежание больших потерь тепла кровь в параллельно расположенных артериях и венах конечностей течет таким образом, что теплая артериальная кровь согревает охлажденную венозную кровь, движущуюся к сердцу (рис. 54, А). Функциональное значение различных отделов петли нефрона неоднозначно. Поступающая из проксимального канальца, в тонкий нисходящий отдел петли нефрона жидкость попадает в зону почки, в интерстициальной ткани которой концентрация осмотически активных веществ выше, чем в корковом веществе почки. Это повышение осмоляльной концентрации в наружной зоне мозгового вещества обусловлено деятельностью толстого восходящего отдела петли нефрона. Его стенка непроницаема для воды, а клетки транспортируют Cl-, Na+ в интерстициальную ткань. Стенка нисходящего отдела петли проницаема для воды. Вода всасывается из просвета канальца в окружающую интерстициальной ткань по осмотическому градиенту, а осмотически активные вещества остаются в просвете канальца. Концентрация осмотически активных веществ в жидкости, поступающей из восходящего отдела петли в начальные отделы дистантного извитого канальца, составляет уже около 200 мосмоль/кг Н2О, т. е. она ниже, чем в ультрафильтрате. Поступление Сl- и Na+ в интерстициальную ткань мозгового вещества увеличивает концентрацию осмотически активных веществ (осмоляльную концентрацию) межклеточной жидкости в этой зоне почки. На такую же величину растет и осмоляльная концентрация жидкости, находящейся в просвете нисходящего отдела петли. Это обусловлено тем, что через водопроницаемую стенку нисходящего отдела петли нефрона в интерстициальную ткань по осмотическому градиенту переходит вода, в то же время осмотически активные вещества остаются в просвете этого канальца. 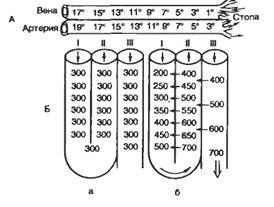 Рис. 54 Функционирование поворотно-противоточной системы (схема).А — теплообменник в сосудистой системе конечностей арктических животных; Б — модель поворотно-противоточной множительной системы в исходном состоянии (а) и в период эффективного концентрирования мочи (б). Чем дальше от коркового вещества по длиннику почечного сосочка находится жидкость в нисходящем колене петли, тем выше ее осмоляльная концентрация. Таким образом, в каждых соседних участках нисходящего отдела петли имеется лишь небольшое нарастание осмотического давления, но вдоль мозгового вещества почки осмоляльная концентрация жидкости в просвете канальца и в интерстициальной ткани постепенно растет от 300 до 1450 мосмоль/кгН2О. На вершине мозгового вещества почки осмоляльная концентрация жидкости в петле нефрона возрастает в несколько раз, а ее объем уменьшается. При дальнейшем движении жидкости по восходящему отделу петли нефрона, особенно в толстом восходящем отделе петли, продолжается реабсорбция Сl- и Na+, вода же остается в просвете канальца. В начальные отделы дистального извитого канальца всегда — и при водном диурезе, и при антидиурезе — поступает гипотоническая жидкость, концентрация осмотически активных веществ в которой менее 200 мосмоль/кг Н2О. При уменьшении мочеотделения вызванном секрецией антидиуретического гормона (АДГ) нейрогипофизом при дефиците воды в организме, увеличивается проницаемость стенки конечных частей дистального сегмента и собирательных трубок для воды. Из гипотонической жидкости, находящейся в связующем канальце и собирательной трубке коркового вещества почки, вода реабсорбируется по осмотическому градиенту, осмоляльная концентрация жидкости в этом отделе увеличивается до 300 мосмоль/кг Н2О, т. е. становится изоосмотичной крови в системном кровотоке и межклеточной жидкости коркового вещества почки. Концентрирование мочи продолжается в собирательных трубках; они проходят параллельно канальцам петли нефрона через мозговое вещество почки. При антидиурезе АДГ увеличивает проницаемость собирательных трубок мозгового вещества почки не только для воды, но и для мочевины, поэтому очищение от мочевины уменьшается при снижении мочеотделения. Прямые сосуды мозгового вещества почки, подобно канальцам петли нефрона, образуют противоточную систему. Благодаря такому расположению прямых сосудов обеспечивается эффективное кровоснабжение мозгового вещества почки, но не происходит вымывания из крови осмотически активных веществ, поскольку при прохождении крови по прямым сосудам наблюдаются такие же изменения ее осмотической концентрации, как и в тонком нисходящем отделе петли нефрона. При движении крови по направлению к вершине мозгового вещества концентрация осмотически активных веществ в ней постепенно возрастает, а во время обратного движения крови к корковому веществу соли и другие вещества, диффундирующие через сосудистую стенку, переходят в интерстициальную ткань. Тем самым сохраняется градиент концентрации осмотически активных веществ внутри почки, и прямые сосуды функционируют как противоточная система. Скорость движения крови по прямым сосудам определяет количество удаляемых из мозгового вещества солей и мочевины и отток реабсорбируемой воды. В случае водного диуреза функции почек отличаются от описанной ранее картины. Проксимальная реабсорбция не изменяется, в дистальный сегмент нефрона поступает такое же количество жидкости, как и при антидиурезе. Осмоляльность мозгового вещества почки при водном диурезе в три раза меньше, чем на максимуме антидиуреза, а осмотическая концентрация жидкости, поступающей в дистальный сегмент нефрона, такая же — приблизительно 200 мосмоль/кг Н2О. При водном диурезе стенка конечных отделов почечных канальцев остается водопроницаемой, а из протекающей мочи клетки продолжают реабсорбировать Na+. В итоге выделяется гипотоническая моча, концентрация осмотически активных веществ в которой может снижаться до 50 мосмоль/кг Н2О. Проницаемость канальцев для мочевины низкая, поэтому мочевина экскретируется с мочой, не накапливаясь в мозговом веществе почки. Таким образом, деятельность петли нефрона, конечных частей дистального сегмента и собирательных трубок обеспечивает способность почек вырабатывать большие объемы разведенной (гипотонической) мочи — до 900 мл/ч, а при дефиците воды экскретировать всего 10—12 мл/ч мочи, в 4,5 раза более осмотически концентрированной, чем кровь. Способность почки осмотически концентрировать мочу исключительно развита у некоторых пустынных грызунов, что позволяет им длительное время обходиться без воды. Количество, состав и свойства мочи. С мочой могут выделяться большинство веществ, имеющихся в плазме крови, а также некоторые соединения, синтезируемые в почке. Почки служат главным органом экскреции конечных продуктов азотистого обмена. У человека при распаде белков образуется мочевина, составляющая до 90 % азота мочи; ее суточная экскреция достигает 25—35 г. С мочой выделяется 0,4—1,2 г азота аммиака, 0,7 г мочевой кислоты (при потреблении пищи, богатой пуринами, выделение возрастает до 2—3 г). Креатин, образующийся в мышцах из фосфокреатина, переходит в креатинин; его выделяется около 1,5 г в сутки. В небольшом количестве в мочу поступают некоторые производные продуктов гниения белков в кишечнике — индол, скатол, фенол, которые в основном обезвреживаются в печени. Белки в нормальной моче выявляются в очень небольшом количестве (суточная экскреция не превышает 125 мг). Небольшая протеинурия наблюдается у здоровых людей после тяжелой физической нагрузки и исчезает после отдыха. Глюкоза в моче в обычных условиях не выявляется. При избыточном потреблении сахара, когда концентрация глюкозы в плазме крови превышает 10 ммоль/л, при гипергликемии иного происхождения наблюдается глюкозурия — выделение глюкозы с мочой. Цвет мочи зависит от величины диуреза и уровня экскреции пигментов. Цвет меняется от светло-желтого до оранжевого. Пигменты образуются из билирубина желчи в кишечнике, где билирубин превращается в уробилин и урохром, которые частично всасываются в кишечнике и затем выделяются почками. С мочой выделяются различные биологически активные вещества и продукты их превращения, по которым в известной степени можно судить о функции некоторых желез внутренней секреции. В моче обнаружены производные гормонов коркового вещества надпочечников, эстрогены, АДГ, витамины (аскорбиновая кислота, тиамин), ферменты (амилаза, липаза, трансаминаза и др.). При патологии в моче обнаруживаются вещества, обычно в ней не выявляемые, — ацетон, желчные кислоты, гемоглобин и др. 31. 2. Невыделительные функции почек. Роль почек в поддержании физиологических констант: осмотического давления, рН, объема жидкости в организме, содержания электролитов, артериального давления. Регуляция деятельности почек. Почки кроме экскреторной функции выполняют и другие функции, являясь непременным исполнительным органом многих гомеостатических функциональных систем. Гомеостатические функции почек. Для поддержания почками постоянства объема и состава внутренней среды и прежде всего крови существуют специальные системы рефлекторной регуляции, включающие специфические рецепторы, афферентные пути и нервные центры, где происходит переработка информации. Команды к почке поступают по эфферентным нервам или гуморальным путем. В целом перестройка работы почки, ее приспособление к непрестанно изменяющимся условиям определяются преимущественно влиянием на гломерулярный и канальцевый аппарат аргинин-вазопрессина [антидиуретического гормона (АДГ)], альдостерона, паратгормона и ряда других гормонов. Одной из важнейших гомеостатических функциональных систем, в деятельности которой почка играет ведущую роль, является функциональная система поддержания водно-солевого баланса (осмотического давления внутренней среды) - ФСОД). Почки являются основным органом осморегуляции. Они обеспечивают выделение избытка воды из организма в виде гипотонической мочи при увеличенном содержании воды (гипергидратация) или экономят воду и экскретируют мочу, гипертоническую по отношению к крови, при обезвоживании организма (дегидратация). При нормальной жизнедеятельности осмолярность межклеточной жидкости может отклоняться в обе стороны от изотоничности. Она повышается при недостаточном поступлении воды в организм, поскольку потеря воды с потом и через легкие превышает выведение осмотически активных веществ. Т.е. гипертоничность внутренней среды вызывается недостатком воды. Реакция организма на недостаток воды совершается следующим образом: усиливается секреция АДГ из нейрогипофиза и возникает чувство жажды. В результате диурез снижается, а возникшее чувство жажды приводит к потреблению воды, что в совокупности восстанавливает осмолярность. 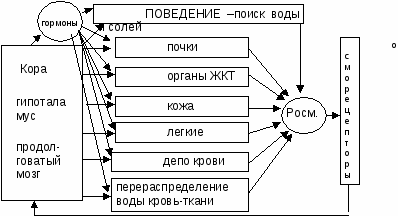  Рис. 55. Структура ФС поддержания осмотического давления После питья воды или при ее избытке в организме снижается концентрация растворенных осмотически активных веществ в крови и падает ее осмоляльность. Это уменьшает активность центральных осморецепторов, расположенных в области супраоптического ядра гипоталамуса, а также периферических осморецепторов, имеющихся в печени, почке и других органах, что приводит к снижению секреции АДГ нейрогипофизом и увеличению выделения воды почкой. При обезвоживании организма или введении в сосудистое русло гипертонического раствора NaCl увеличивается концентрация осмотически активных веществ в плазме крови, возбуждаются осморецепторы, усиливается секреция АДГ, возрастает всасывание воды в канальцах, уменьшается мочеотделение и выделяется осмотически концентрированная моча. В эксперименте было показано, что, помимо осморецепторов, секрецию АДГ стимулируют натриорецепторы. При введении в область III желудочка мозга гипертонического раствора NaCl наблюдался антидиурез, если же вводить в то же место гипертонический раствор сахарозы, то мочеотделение не уменьшается. Помимо осмо- и натриорецепторов, уровень секреции АДГ определяет активность волюморецепторов, воспринимающих изменение объема внутрисосудистой и внеклеточной жидкости. Ведущее значение в регуляции секреции АДГ имеют рецепторы, которые реагируют на изменение напряжения сосудистой стенки в области низкого давления. Прежде всего, это рецепторы левого предсердия, импульсы от которых передаются в ЦНС по афферентным волокнам блуждающего нерва. При увеличении кровенаполнения левого предсердия активируются волюморецепторы и угнетается секреция АДГ, что вызывает усиление мочеотделения. Поскольку активация волюморецепторов в отличие от осморецепторов обусловлена увеличением объема жидкости, т. е. возросшим содержанием в организме воды и солей натрия, возбуждение волюморецепторов приводит к увеличению экскреции почкой не только воды, но и натрия. Эти процессы связаны с секрецией натрийуретического гормона, уменьшением секреции ренина, ангиотензина, альдостерона, при этом снижается тонус симпатической нервной системы, в результате уменьшается реабсорбция натрия и возрастают натрийурез и мочеотделение. В конечном счете восстанавливается объем крови и внеклеточной жидкости. Почки в случае необходимости удерживают воду или выделяют лишнюю, способствуя поддержанию необходимого объема крови. Р
оль почек в регуляции ионного состава крови. Почки являются эффекторным органом системы ионного гомеостаза. В организме существуют системы регуляции баланса каждого из ионов. Для некоторых ионов уже описаны специфические рецепторы, например натриорецепторы. Рефлекторная регуляция транспорта ионов в почечных канальцах осуществляется как периферическими, так и центральными нервными механизмами. Регуляция реабсорбции и секреции ионов в почечных канальцах осуществляется несколькими гормонами. Реабсорбция натрия возрастает в конечных частях дистального сегмента нефрона и собирательных трубочках под влиянием гормона коркового вещества надпочечника альдостерона. Этот гормон выделяется в кровь при уменьшении концентрации натрия в плазме крови и уменьшении объема циркулирующей крови. В усилении выделения натрия почкой участвует натрийуретический гормон, одним из мест образования которого является предсердие. При увеличении объема циркулирующей крови, повышении объема внеклеточной жидкости в организме усиливается секреция в кровь этого пептидного гормона. Секрецию калия в дистальном сегменте и собирательных трубочках усиливает альдостерон. Инсулин уменьшает выделение калия. Алкалоз сопровождается усилением выделения калия, а при ацидозе калийурез уменьшается. При уменьшении концентрации кальция в крови паращитовидные железы выделяют паратгормон, который способствует нормализации уровня кальция в крови, в частности благодаря увеличению его реабсорбции в почечных канальцах и высвобождению из кости. При гиперкальциемии, а также под влиянием гастрина (или подобного ему вещества), вырабатываемого в пищеварительном тракте в процессе всасывания кальция, стимулируется выделение в кровь клетками щитовидной железы кальцитонина, который способствует уменьшению концентрации Са2+ в плазме крови благодаря увеличению экскреции почкой и переходу Са2+ в кость. В регуляции обмена Са2+ участвуют образующиеся в почке активные формы витамина Dз. В почечных канальцах регулируется уровень реабсорбции Mg2+, Cl-, SO42-, а также микроэлементов. Р
оль почек в регуляции кислотно-основного состояния. Являясь важным исполнительным органов функциональной системы поддержания рН, почки участвуют в поддержании постоянства концентрации Н+ в крови, экскретируя кислые продукты обмена. Активная реакция мочи у человека и животных может очень резко меняться в зависимости от состояния кислотно-основного состояния организма. Концентрация Н+ при ацидозе и алкалозе различается почти в 1000 раз, при ацидозе рН может снижаться до 4,5, при алкалозе — достигать 8,0. Это способствует участию почек в стабилизации рН плазмы крови на уровне 7,36. При питании мясом образуется большее количество кислот, и моча становится кислой, а при потреблении растительной пищи рН сдвигается в щелочную сторону. При интенсивной физической работе из мышц в кровь поступает значительное количество молочной и фосфорной кислот и почки увеличивают выделение «кислых» продуктов с мочой. Кислотовыделительная функция почек во многом зависит от кислотно-основного состояния организма. Так, при гиповентиляции легких происходит задержка СО2 и снижается рН крови — развивается дыхательный ацидоз, при гипервентиляции уменьшается напряжение СО2 в крови, растет рН крови — возникает состояние дыхательного алкалоза. Содержание ацетоуксусной и β-оксимасляной кислот может нарастать при нелеченом сахарном диабете. В этом случае резко снижается концентрация гидрокарбоната в крови, развивается состояние метаболического ацидоза. Рвота, сопровождающаяся потерей соляной кислоты, приводит к увеличению в крови концентрации гидрокарбоната, что приводит к метаболическому алкалозу. В конечном счете почки стабилизируют концентрацию гидрокарбоната в плазме крови на уровне 26—28 ммоль/л, а рН — на уровне 7,36. Участие почек в регуляции системного артериального давления. При снижении кислородного снабжения ткани почек, клетки ЮГА вырабатывают гормон ренин, который через систему ангиотензин-альдостерон влияет на системное кровяное давление и кровоток в почках. Инкреторная функция почек. В почках вырабатывается несколько биологически активных веществ, позволяющих рассматривать ее как инкреторный орган. Гранулярные клетки юкстагломерулярного аппарата выделяют в кровь ренин при уменьшении артериального давления в почке, снижении содержания натрия в организме. В почке синтезируется активатор плазминогена — урокиназа. В мозговом веществе почки образуются простагландины. Они участвуют, в частности, в регуляции почечного и общего кровотока, увеличивают выделение натрия с мочой, уменьшают чувствительность клеток канальцев к АДГ. Клетки почки извлекают из плазмы крови образующийся в печени прогормон — витамин D3 и превращают его в физиологически активный гормон — активные формы витамина D3. Этот стероид стимулирует образование кальцийсвязывающего белка в кишечнике, способствует освобождению кальция из костей, регулирует его реабсорбцию в почечных канальцах. Почка является местом продукции эритропоэтина, стимулирующего эритропоэз в костном мозге. В почке вырабатывается брадикинин, являющийся сильным вазодилататором. Метаболическая функция почек. Данная функция обусловлена участием почек в обеспечении постоянства концентрации в крови ряда физиологически значимых органических веществ. Почки участвуют в обмене белков, липидов и углеводов. В почечных клубочках фильтруются низкомолекулярные белки, пептиды. Клетки проксимального отдела нефрона расщепляют их до аминокислот или дипептидов и транспортируют через базальную плазматическую мембрану в кровь. Это способствует восстановлению в организме фонда аминокислот, что важно при дефиците белков в рационе. При заболеваниях почек эта функция может нарушаться. Почки способны синтезировать глюкозу (глюконеогенез). При длительном голодании почки могут синтезировать до 50 % от общего количества глюкозы, образующейся в организме и поступающей в кровь. Для энерготрат почки могут использовать глюкозу или свободные жирные кислоты. При низком уровне глюкозы в крови клетки почки в большей степени расходуют жирные кислоты, при гипергликемии преимущественно расщепляется глюкоза. |