Курс лекций по биомеханике. Курс лекций Содержание Биомеханика как учебная и научная
 Скачать 5.94 Mb. Скачать 5.94 Mb.
|

|
2. СОЕДИНЕНИЯ ЗВЕНЬЕВ Соединения звеньев в биокинематических цепях обусловливают многообразие возможностей движении. От способа соединения и участия мышц в движениях зависит их направление и размах (пространственная. форма движений). 2.1. Кинематические пары Кинематическая пара — это подвижное соединение двух звеньев. Способ соединения накладывает ограничения (связи) на относительное движение (степени связи); наличие подвижности в соединении предоставляет звеньям определенные возможности относительного движения (степени свободы движения). Различают связи: а) геометрические (постоянные препятствия перемещению в каком-либо направлении) и б) кинематические (ограничение скорости, например, мышцей-антагонистом) . Кинематические пары бывают: а) поступательные — одно звено может перемещаться поступательно по другому (например, боковые движения нижней челюсти); б) вращательные (па-пример, повороты в наиболее распространенных в теле человека цилиндрических и шаровидных суставах); в) винтовые с сочетанием поступательного и вращательного движений (например, в голеностопном суставе). Соединения, допускающие поворот звеньев пары, называют шарнирами. 2.2. Кинематические цепи Кинематическая цепь — это последовательное или разветвленное соединение ряда кинематических пар. Кинематическую цепь, в которой конечное звено свободно, называют незамкнутой, а цепь, в которой нет свободного конечного звена,— замкнутой. В каждом соединении незамкнутой цепи возможны изолированные движения. Они геометрически независимы от движений в других соединениях (если не учитывать взаимодействия мыши). Например, свободные конечности, когда их концевые звенья свободны (рис. 4, а), представляют незамкнутые цепи. Замкнутымикинематическими цепями в теле человека являются, например, грудина, ребро, позвоночник, ребро и снова грудина. Такие замкнутые цепи разомкнуть невозможно. Незамкнутые могут замыкаться, причем часто через опору (см. рис. 4, а). В сложной пирамиде, составленной несколькими акробатами, образуются даже своего рода «сети» (в плоскости) и «решетки» (в пространстве) с очень сложной взаимной зависимостью движений звеньев. В замкнутой или замкнувшейся цепи невозможно одиночное изолированное движение, т. е. движение в одном соединении. Так, сгибая и выпрямляя ноги в выпаде, можно убедиться в том, что движение в любом суставе непременно вызывает движения и в других (см. рис. 4, б). Таким образом, движения в незамкнутых цепях характеризуются относительной независимостью звеньев. В замкнутых же, а также замкнувшихся цепях движения одних звеньев влияют на движения даже отдаленных звеньев (помогают или мешают). Например, сгибая руки в локтевых суставах в упоре лежа (замкнувшаяся цепь), можно или отводить плечи, или разгибать их в плечевых суставах. Тогда будут растягиваться либо приводящие мышцы плеча (большие грудные, широчайшие спины и др.), либо сгибатели (передние части дельтовидных). Теперь при разгибании рук в локтевых суставах приводящие мышцы или сгибатели плечевых суставов в зависимости от того, какие мышцы растянуты, могут передавать свое действие на локтевые суставы — способствовать разгибанию рук в этих суставах. Передача действия мышц на смежные и отдаленные суставы характерна для замкнутых цепей и часто имеет место в подобных условиях работы мышц. В замкнутых цепях возможностей движений меньше, но управление ими точнее, чем в незамкнутых. 2.3. Степени свободы движений Число степеней свободы движений соответствует количеству возможных независимых линейных и угловых перемещений тела. Тело, ничем не ограниченное в движениях (может двигаться в любом направлении), называется свободным. Движение свободного тела возможно в трех основных направлениях — вдоль осей координат, а также вокруг этих трех осей; оно имеет 6 степеней свободы движения (рис. 5, а). Наложение связей уменьшает количество степеней свободы (табл. 1). Если закрепить одну точку тела, то сразу снимается 3 степени свободы: тело не сможет двигаться вдоль трех осей координат; у него останутся только возможности вращения вокруг этих осей, т. е. только три степени свободы (см. рис. 5, б). Так соединены кости трехосных (шаровидных) с у с т а в о в. 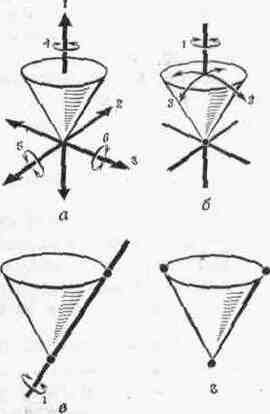 При закреплении двух точек в теле возможно вращение лишь вокруг линии (оси), проходящей через обе точки (см. рис. 5, в). Так соединены кости одноосных суставов, обеспечивающих одну степень свободы. Если же закреплены три точки (не лежащие на одной линии), то движения тела совсем невозможны (см. рис. 5, г). Та кое соединение неподвижно и, следовательно, не является суставом. Как известно, двуосные суставы (эллипсовидный — лучезапястный, седловидный — первый запястно-пястный) обеспечивают вторую степень свободы благодаря неполному взаимному соответствию своих суставных поверхностей (неконгруэнтность). По этой же причине, если рука в локтевом суставе согнута, возможны приведение и отведение локтевой кости в плечелоктевом сочленении (например, при поворотах отвертки, штопора, ключа вокруг оси, проходящей вдоль второй пястной кости. в преобладающей части суставов тела человека 2 или 3 степени свободы. При нескольких степенях свободы движений (двух и более) возможно бесчисленное множество траекторий. Значит, в движениях в неодноосных суставах отсутствует определенность, задаваемая способом соединения. Тем более это характерно для цепей с несколькими неодноосными суставами. Множество возможностей движений в суставах кинематической пары более чем с одной степенью свободы в отличие от технической пары требует для выполнения каждого определенного движения: а) выбор необходимой траектории, б) управления движением по траектории (направлением и величиной скорости) и в) регуляции движения, понимаемой как борьба с помехами, сбивающими с траектории (см. гл. IV). 2.4. Геометрия движений Число основных осей сустава соответствует количеству степеней свободы движений одного звена относительно другого. Плоскость движения перпендикулярна оси вращения и характеризует направление перемещения звена. Размах движений — это угловое перемещение звена из одного крайнего положения в другое. В суставном движении различают ось, плоскость и размах. Все пары звеньев связаны в суставах неразрывно, как в шарнирах, поэтому они могут двигаться в основном только вокруг осей (не считая незначительного скольжения). Однако не существует суставов совершенно правильной геометрической формы. А если бы даже такие и были, то при сдавливании гиалиновых суставных хрящей форма суставных поверхностей нарушалась бы. Следовательно, геометрические оси вращения не постоянны и правильнее говорить о мгновенных осях вращения. В связи с этим количество осей в суставе означает в биомеханическом смысле только количество степеней свободы движения, а не постоянных геометрических осей вращения. Продольные оси звеньев чаще всего не строго перпендикулярны геометрическим осям вращения. Поэтому различные точки звеньев движутся в плоскостях, параллельных друг другу, а сама продольная ось звена описывает поверхности, близкие к коническим. Таким образом, плоскость движения в суставе характеризует, в какой плоскости движутся точки звена. Она перпендикулярна геометрической оси вращения и не обязательно совпадает с плоскостью движения продольной оси самого звена. Размахом движений измеряется подвижностьв каждом отдельном суставе1. Наибольший размах бывает в пассивных движениях; с увеличением внешней нагрузки (вес отягощения, силы инерции) растягиваются мягкие ткани-ограничители. Размах же в активном движении меньше, чем в пассивном, так как там мышцы имеют предел силы, в то время как величина внешних сил не ограничена. К тому же сила собственных мышц пары звеньев приложена невыгодно при крайних положениях звеньев в суставе. Подвижностью соседних звеньев в соединениях каждой пары обусловлена гибкость всей кинематической цепи в целом. Гибкость цепи (например, позвоночника) измеряется общим размахом движения концевого звена относительно другого конца цепи. Общий размах движения в целом бывает меньше суммы размахов изолированных движений в суставах, так как вследствие пассивной недостаточности многосуставных мышц возникают дополнительные связи. 3. ЗВЕНЬЯ КАК РЫЧАГИ Скелет, составленный из подвижно соединенных костей, представляет собой твердую основу биокинематических цепей. Звенья цепей с приложенными к ним силами (мышечной тяги и др.) в биомеханике рассматриваются как система составных рычагов. 3.1. Виды рычагов в теле человека Рычаг — твердое тело, которое может под действием приложенных сил вращаться вокруг опоры (оси) в двух противоположных направлениях, а также сохранять свое положение. Как простейший механизм, рычаг служит для передачи движения и силы на расстояние. Каждый рычаг имеет точку опоры — ось рычага (О на рис. 6), точки (А, В) приложения двух взаимно противодействующих сил (G и Q). Чтобы определить расстояние до места приложения сил от оси рычага, измеряют плечи рычага (О А и 0В). Чтобы определить расстояния до направления действия сил от оси рычага, измеряют плечи сил {а. и k — перпендикуляры, опущенные из точки О на линии действия сил, т. е. их направления). Рычаги бывают о д н о п л еч и е (с точкой опоры на конце рычага — см. рис. 6,а)и двуплечие (с точкой опоры между концами рычага — см. рис. 6, б). Принципиальной разницы в отношении работы сил между ними нет: поворачивая плечо двуплечего рычага вокруг точки О из положения do в di, затем da, можно перейти к рычагу одноплечему (см. рис. 6, б). У нового, одноплечего, рычага иная форма, но те же свойства, что и у прежнего, двуплечего, рычага. Звенья тела человека имеют в своей основе костные рычаги. Двуплечие рычаги используются, например, при сохранении положения стоя, а одноплечие — в быстрых движениях конечностей. Одно и то же звено для разных мышц может быть то одноплечим, то двуплечим рычагом. 3.2. Условия сохранения положения звеньев и их движения как рычагов Для равновесия рычага необходимо равенство моментов приложенных сил противоположного действия относительно оси рычага; для ускорения рычага — неравенство этих моментов сил. В результате действия противоположных сил звено как рычаг может: а) сохранять положение или продолжать движение с прежней скоростью и б) получить ускорение в сторону той или иной силы. Эффект совместного действия сил зависит от соотношения их моментов. Если моменты обеих сил равны, то либо сохраняется неподвижное положение рычага, либо продолжается движение со скоростью, имевшейся в момент уравнивания обоих моментов. Когда момент одной из сил больше момента другой силы, возникает ускорение в ту сторону, куда направлена сила большего момента. Если больше момент силы мышечной тяги, мышца сокращается (преодолевающая работ а) и плечо рычага передвигается в ее сторону. Если больше момент силы отягощения, то мышца растягивается (уступающая работ а) и плечо рычага передвигается в другую сторону. Следует заметить, что в рассмотренных случаях рычаг для упрощения расчета считается невесомым и безынерционным. В действительности же звенья тела человека обладают весом и при ускорениях оказывают инертное противодействие в зависимости от своих моментов инерции. В описанном выше примере (см. рис. 6, а) рычаг расположен горизонтально, поэтому направление веса отягощения перпендикулярно рычагу и плечо силы совпадает с плечом рычага (k). В движениях человека это бывает редко; чаще всего силы приложены под острым или тупым углом. Тогда плечо силы меньше плеча рычага и, следовательно, момент силы меньше максимально возможного для данной величины силы. При тупом или остром угле приложения силы F (см. рис. 6, д, е) плечо силы (d) — катет, а плечо рычага (/) — гипотенуза в прямоугольном треугольнике. Следовательно, плечо силы меньше максимального (см. рис. 6, г, где /—d), и момент силы поэтому тоже не наибольший. Это же видно при разложении сил: нормальная составляющая (Fpf), перпендикулярная направлению движения, только прижимает рычаг к точке опоры или оттягивает от нее и непосредственно не влияет на скорость движения. Тангенциальная же составляющая (Рт), касательная к траектории конца рычага, влияет на скорость движения. Поэтому тангенциальная тяга называется вращающей (или явной), а нормальная — укрепляющей (или скрытой). При движениях изменяются положения костных рычагов и углы приложения сил. Следовательно, изменяются плечи сил. Длина мышц тоже не остается постоянной, а поэтому и их напряжения становятся больше или меньше. Таким образом, моменты сил, их соотношение, а отсюда и условия сохранения положения или движения частей тела как рычагов не постоянны. 3.3. «3олотое правило» механики1 в движениях человека Работа, совершаемая силою, приложенной на одном плече рычага, передается на другое плечо. Сила тяги мышцы, приложенная на коротком плече рычага, вызывает во столько раз большее смещение другого плеча, во сколько первое плечо короче второго; налицо выигрыш в пути. В связи с тем, что разные пути проходятся за одно и то же время, здесь имеется выигрыш в скорости. Сила, передаваемая на длинное плечо рычага, как раз во столько же раз меньше, чем приложенная. Таким образом, выигрыш в скорости достигается за счёт проигрыша в силе. Почти все мышцы в теле человека прикрепляются вблизи суставов (короткое плечо рычага); это приводит к выигрышу в пути (а, следовательно, и в скорости) при проигрыше в силе. При большей части положений костного рычага мышечные тяги направлены под острым или тупым углом к звену (вдоль звена), что влечет невосполнимые потери в силе мышц (уменьшается вращающая тяга). Нормальная же (скрытая) тяга в этом случае способствует укреплению сустава, через который мышца переходит. При больших нагрузках напрягаются все мышцы, окружающие сустав, в том числе и антагонисты. При этом резко возрастают потери в суммарной тяге мышц; в то же время достигается и положительный эффект — укрепление нагруженного сустава. В связи с особенностями приложения мышечных тяг к костным рычагам необходимы весьма значительные напряжения мышц для выполнения не только силовых, но и скоростных д в и ж е н и и. Входящие в биокинематические цепи звенья тела образуют системы составных рычагов, в которых «золотое правило» механики проявляется намного сложнее, чем в простых одиночных рычагах. 4. БИОМЕХАНИЧЕСКИЕ СВОЙСТВА МЫШЦ Мышцы как физические тела обладают рядом механических свойств — упругостью, вязкостью, ползучестью, релаксацией. Как биологические объекты мышцы проявляют свойства возбудимости и сократимости. Все названные свойства тесно взаимосвязаны, что важно учитывать при биомеханическом исследовании движений. 4.1. Механические свойства мышц Упругость проявляется в возникновении напряжения в мышце при ее деформации под действием нагрузки. Вязкость — в замедлении деформации внутренними силами (жидким трением, молекулярными силами). На графике «длина — напряжение», полученном на изолированной мышце экспериментально (рис. 7, а), удобно рассмотреть упругостьмышцы. Здесь видно, как по мере увеличения нагрузки мышца удлиняется и как при этом растет ее напряжение. Отсюда следует: 1) нагрузка (Р) растягивает мышцу, удлиняя ее (на l), т. е. для растягивания мышцы необходимо приложить силу; 2) по мере удлинения мышцы (на l) ее напряжение увеличивается (на F); следовательно, чтобы вызвать напряжение мышцы (без дополнительного возбуждения), необходимо ее растянуть; 3) приложенная нагрузка (Р) определяет величину напряжения мышцы (F); таким образом, чтобы получить большое напряжение, надо приложить большую нагрузку (сопротивление тяге мышцы) — действие равно противодействию; 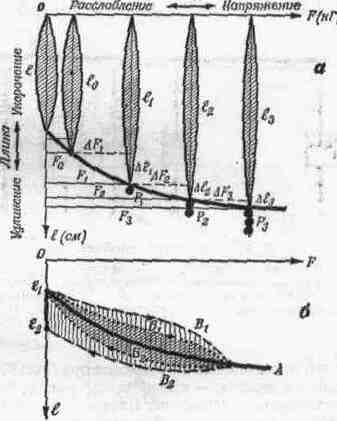 Рис.7. Графики «длина—напряжение» мышцы: а — проявление упругости (зависимость деформации и напряжения от нагрузки); б — проявление вязкости («петли гистерезиса») (ориг.) 4) упругость мышцы нелинейна ( l2 больше l3 при равных F2 и F3); следовательно, по мере значительного растягивания одинаковые приращения длины мышцы дают все большие приращения напряжения; 5) при отсутствии нагрузки длина мышцы (l) является наименьшей («свободная длина» мышцы) — нерастянутая мышца не напряжена; 6) в условиях организма длина мышцы (/о) больше «свободной длины» и мышца несколько напряжена, т. е. всегда обладает «тонусом» покоя (F0). Таковы особенности упругих свойств невозбужденной мышцы. Вязкость мышцы как причину запаздывания деформации можно наблюдать по отклонению графика действительной деформации от линии зависимости «длина — напряжение» в установившемся состоянии (см. рис. 7, б— линия A). При меньшей вязкости (линии Б) изменение длины отстает от изменения напряжения как при растягивании мышцы (£i), так и при ее сокращении (Бz). В этом случае мышца, хотя и не сразу, но все же возвратилась к исходному состоянию (точка l3). При большей вязкости (линии В) замедление еще больше и мышца не возвратилась к прежнему состоянию (точка l3) — обнаружилась остаточная деформация (расстояние l3 — l1). Полученные кривые (Б, В) образуют так называемые «петли гистерезиса», характеризующие запаздывание процесса деформации из-за тормозящего действия вязкости. При этом неизбежна потеря энергии. Она пропорциональна площади, ограниченной петлей гистерезиса (косая штриховка для Б1—Бz и вертикальная—для B1—Вz)- Действительно, площадь, определяемая произведением силы (напряжение) и пути (изменение длины), равна работе, затраченной на преодоление вязкости. Ползучесть — это свойство мышцы изменять с течением времени соотношение «длина — напряжение»: нагруженная (напряженная) мышца имеет соответствующую длину; через некоторое время при тех же нагрузках и напряжении эта длина увеличивается. Релаксациязаключается в том, что растянутая мышца, сохраняя длину, постепенно с течением времени уменьшает свое напряжение, расслабляется. Совокупность этих механических свойств (упруговязких, ползучести и релаксации) во всевозможных сочетаниях в различных условиях, в сущности, и есть то, что называется эластичностью1мышцы. Высокоэластичноймышце свойственны значительная растяжимость, большая жесткость при большом растягивании (нелинейная упругость) и малые потери энергии (небольшая вязкость) при деформациях. И хотя механизм, обеспечивающий названные свойства, еще неполностью объяснен, их проявления очень важно учитывать при изучении способов повышения эффективности действия мышц в движениях. 4.2. Режимы работы мышц Режим работы мышцы определяется изменением либо ее длины, либо ее напряжения, либо того и другого одновременно. Возбудимость мышцы проявляется в изменении как ее напряжения, так и механических свойств — упругости, вязкости и др. В результате возбуждения химическая энергия в мышце превращается в механическую. Возбужденная мышца при той же нагрузке и напряжении имеет меньшую длину — не меняя своего напряжения, она сокращается. 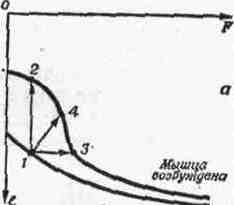 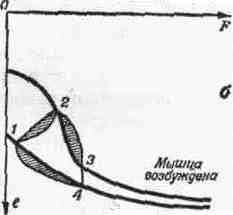 Мышца не возбуждена Мышца не возбуждена Рис. 9. Работа мышцы: а — проявление возбуждения: режимы — изотонический (1—2), изометрический (1—3), ауксотонический (1—4); б — проявление вязкости мышцы (орг.) Свойство сократимости удобно разобрать на графиках «длина — напряжение» невозбужденной и максимально возбужденной мышц (рис. 9, а). Вся кривая, относящаяся к максимально возбужденной мышце, расположена выше, чем кривая покоющейся. Следовательно, при равной величине напряжения длина возбужденной мышцы меньше. Кривые, полученные при разных степенях возбуждения мышц, занимают промежуточное положение между упомянутыми крайними случаями. Можно этот же график разобрать иначе: точки кривой максимально возбужденной мышцы расположены правее (например, точки1и 3) точек графика покоющейся мышцы, имеющей ту же длину. Значит, при одной и той же длине мышца, будучи возбужденной, напряжена больше, чем в состоянии покоя. Если в подготовительном движении (например, замах, подседание) мышца перед сокращением растягивается, то она тормозит движение; кинетическая энергия тормозимого звена переходит в потенциальную энергию упругой деформации мышцы. Теперь растянутая мышца напряжена; в ней накоплена потенциальная энергия упругой деформации. С началом активного движения при возбуждении мышцы в ней образуется механическая энергия напряжения, освобождаемая при биохимической реакции. Вся биопотенциальная энергия мышцы состоит из превращенной биохимической и упругой механической. Преобразование всей биопотенциальной энергии мышцы в механическую (кинетическую) по-разному проявляется в различных условиях работы мышцы. Линия перехода па графике от точки 1 к точке 2 изображает сокращение мышцы в изотоническом режиме (без изменения напряжения — см. рис. 9, а). В реальных движениях в живом организме такой режим вряд ли может встретиться. При движениях изменяются моменты сил мышечной тяги, а также других сил, поскольку изменяются углы их приложения, а у мышц — и их длина. Сохранять величину напряжения мышцы в этих условиях практически невозможно, да это и не нужно. Линия перехода на графике от точки 1 к точке 3 изображает увеличение напряжения мышцы при ее работе в изометрическом режиме. Например, при непреодолимых препятствиях длина мышцы не изменяется, однако напряжение ее вследствие возбуждения увеличивается. Это случай «статической работы» мышц при сохранении положения тела (см. гл. VI). Для работы мышц человека при движениях обычен так называемый ауксотонический режим1(например, линия перехода на графиках от точки 1 к точке 4),связанный с изменением и длины, и напряжения мышцы. Этот режим правильнее было бы называть «ауксоническим», учитывая, что изменяется не только напряжение, но и длина мышцы. На графиках реальных движений все рассмотренные переходы происходят не по прямым линиям, так как вязкость замедляет деформацию. На рис. 9, б представлены петли гистерезиса при возбуждении ранее нагруженной мышцы (/—2), при дополнительной нагрузке возбужденной мышцы {2—3) и после снятия возбуждения при ее разгрузке {4—I}. Заштрихованные участки соответствуют потерям энергии на преодоление вязкости. Считают, что вязкость мышц увеличивается при быстрых движениях и при значительном возбуждении, т. е. как раз в условиях соревновательной борьбы спортсмена. Однако разогревание мышц при разминке снижает вязкость, уменьшает торможение при сокращении и растягивании мышц. Следовательно, на соревнованиях и тренировках важно для снижения вязкости сохранять в разогретых мышцах тепло. 5. МЕХАНИЧЕСКОЕ ДЕЙСТВИЕ МЫШЦ Механическое действие мышц проявляется в основном как тяга. Мышечная тяга характеризует величину приложенной силы мышц и ее направление — она образуется при суммировании сил тяги всех ее волокон. 5.1. Величина и направление тяги мышцы Тяга мышцы зависит от совокупности механических, анатомических и физиологических условий. К механическим условиям относится нагрузка — как растягивающая мышцу, так и противодействующая ее сокращению. С увел и ч е н и е м длины растягиваемой мышцы растет ее упругое напряжение (если возбуждение неизменно). Это особенно выражено при больших растягиваниях, в связи с проявлением нелинейной упругости. Сила тяги мышцы нарастает лишь до известного предела увеличения нагрузки, после которого дальнейший рост нагрузки уже не вызывает увеличения силы тяги мышцы. 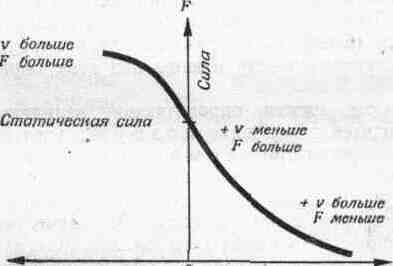 -V Растягивание Сокращение +V Скорость Рис. 10. Зависимость между быстротой сокращения и напряжением мышцы (схема по Абботту и др.) С увеличением преодолеваемой нагрузки (в известных пределах) сила тяги мышцы становится больше, но быстрота сокращения падает (рис. 10). Однако при уступающей работе, выполняемой с ускорением, напряжение мышцы растет. Большая сила тяги может проявится также при большом ускорении тела, имеющего малую массу. В связи с этим от выбора отягощений и режима работы мышц при подборе упражнений для воспитания силовых качеств1 зависит ход развития тех или иных сторон силовой подготовки. К анатомическим условиям проявления тяги мышцы относятся строение мышцы и ее расположение (в данный момент движения). От строения мышцы зависит ее физиологический поперечник, который определяют по сечению, проходящему через все волокна в мышце перпендикулярно их осям. Но дело не только в суммарной силе тяги всех волокон мышцы. От расположения волокон зависит и степень нелинейности упругих свойств. Так, в мышцах с косым ходом волокон при малом растяжении происходит очень большое увеличение упругих сил. Расположение мышцы относительно оси сустава и звена в данный момент движения влияет, во-первых, на величину плеча силы, а стало быть, и величину момента силы тяги. При острых (менее 45°) и тупых (более 135°) углах вращающая тяга меньше укрепляющей. Во-вторых, расположение мышцы влияет на направление тяги мышцы. Напряженная мышца стремится сблизить места прикрепления (центры их площадей) обоих своих концов. Только если брюшко или сухожилие переходит через костный выступ (блок), то направление тяги определяется прямой, соединяющей середину толщи мышцы над этим блоком с местом ее прикрепления. Физиологические условия, определяющие величину тяги мышцы, в основном сводятся к условиям в о з б у ж.д е н и я мышцы и его изменения, в частности при утомлении. Как известно, пучок от 10 до 3000 мышечных волокон (мион) иннервируется одним нервным волокном — отростком одной двигательной нервной клетки передних рогов серого вещества спинного мозга. От количества возбужденных мионов в основном зависит сила тяги мышцы. Максимальное возбуждение наибольшего количества мионов обеспечивает наибольшую силу тяги мышцы. В связи с утомлением существенно изменяется работоспособность мышцы. Это следует учитывать при биомеханическом исследовании спортивной техники. 5.2. Результат тяги мышцы Результат приложения тяги мышцы в кинематической цепи зависит от: а) закрепления звеньев; б) соотношения сил, вызывающих движение, и сил сопротивления, в) начальных условий вращения. Чтобы определить результат тяги мышцы, недостаточно установить величину и направление этой тяги. При различных условиях закрепления звеньев одна и та же тяга приводит к неодинаковому результату — разным движениям звеньев в суставе. В кинематической паре может быть закреплено одно звено, или оба свободны, или оба закреплены (рис. 11). Соответственно возникнут ускорения того или иного звена, либо обоих вместе (встречные движения), либо соединение будет фиксировано. Для двусуставной мышцы, не говоря уже о многосуставных, число возможных вариантов результата во много раз больше. Когда одно или оба звена могут двигаться свободно, величина ускорения зависит от вращающего момента, момента сил внешнего сопротивления и распределения масс звена. Иначе говоря, движение зависит от тог о, каково соотношение движущей силы и соответствующего сопротивления. Наконец, особо важны начальные условия вращения — положение звеньев пары и скорость их движения (направление и величина) в момент приложения силы. От них зависит эффект тяги мышцы: ускорение движения, замедление его или изменение направления, изменение величины и направления скорости одновременно. В кинематических цепях действуют те же факторы, определяющие результат тяги каждой мышцы. Но так как в кинематической цепи все звенья так или иначе взаимосвязаны, то в каждом конкретном случае лишь совокупность всех факторов определяет, результат работы мышц в целом. 5.3. Виды и разновидности работы мыши, В зависимости от изменения длины мышцы различают следующие виды ее работы: а) статическая (изометрический режим)— длина мышцы не изменяется, б) динамическая — мышца либо укорачивается (преодолевающая работа –миометрический режим), либо удлиняется (уступающая работа –плиометрический режим)1. В зависимости от сочетания трех видов работы мышцы (изменение длины) с возможными вариантами изменения ее напряжений может быть девять элементарных (типичных) разновидностей работы мышц . При сохранении положения тела чаще встречается постоянная фиксация (постоянная длина)(разновидность 5). В движениях наиболее обычны разгон (до максимума скорости)(3) и торможение (до остановки) (7). В точных движениях характерно притормаживание (9). Силовая работа выполняется с напряжениями в движениях «до отказа» (1) и усилением фиксации (4). При выполнении одного и того же действия могут иметь место смены (иногда неоднократные) разновидностей работы у одной и той же мышцы. Самая характерная смена работы — с уступающей на преодолевающую. Растягиваемая мышца поглощает кинетическую энергию тормозимого звена, превращая ее в потенциальную энергию упругой деформации и при последующем сокращении — вновь в кинетическую энергию (реверсивный режим— со сменой направления движения). . 6. ГРУППОВЫЕ ВЗАИМОДЕЙСТВИЯ МЫШЦ Мышцы, влияющие на движения биокинематических цепей, как правило, функционируют не изолированно, а группами. Взаимодействие осуществляется между мышцами внутри групп, а также между группами мышц. 6.1. Рабочие и опорные напряжения Рабочие напряжения мышц (динамическая работа) обусловливают движения, а опорные напряжения мышц (статическая работа) создают необходимую для этого опору. Чаще всего человек движется, сохраняя опору. При этом можно отчетливо выделить звенья, имеющие связь с общей опорой тела, т. е. опорные з в е н ь я, и движущиеся относительно опорных—подвижные звенья. Кроме того, многие звенья тела служат опорой для мышц, вызывающих движения других звеньев. Они также делятся на опорные и подвижные звенья. Опорные звенья сохраняют взаимное расположение, а иногда и связь с опорой, благодаря мышцам, развивающим опорные напряжения2.В теле создается опорная цепь («стойка» механизма), как бы фундамент для подвижных звеньев. В этом случае подвижные звенья (рабочая цепь) получают закрепленные точки для мышц, создающих рабочие напряжения. Одна часть мышц непосредственно связана с опорными звеньями; другая — расположена между подвижными звеньями. Мышцы, развивающие опорные напряжения, выполняют статическую работу, а развивающие рабочие напряжения—динамическую. При движениях человека вне опоры цепи звеньев его тела нередко объединяются в блоки благодаря опорным напряжениям мыши. В этих случаях движения происходят не во всех суставах тела. Часто в действиях спортсмена статические опорные напряжения заменяются опорными динамическими напряжениями — с малым размахом движения. Так, туловище при различных видах ходьбы или бега обычно не зафиксировано полностью: почти всегда его опорная функция осуществляется при движениях с малым размахом. Если рабочие напряжения мышц, вызывающих движения подвижных звеньев, велики, то опорные напряжения, закрепляющие неподвижные звенья, тоже должны быть велики. Их напряжения взаимно связаны. 6.2. Взаимодействующие группы мышц Мышцы, окружающие сустав, при движении разделяются на функциональные группы: а) синергисты (совместного действия), выполняющие преодолевающую работу, и б) их антагонисты2 (противоположного действия), выполняющие уступающую работу Мышцы-синергисты (Si и 5з — рис. 12) тянут под углом друг к другу. Одна составляющая тяги у них направлена одинаково (движущая тяга) (D» и Da), а другая—противоположно (Ri и Pi). Противоположно направленные составляющие взаимно нейтрализуются, а одинаково направленные — суммируются. По-особому участвуют в движениях мышцы-антагонисты. Нет такого движения, при котором они не растягивались бы. Будучи растянутыми, они могут вызвать движение звена тела в обратном направлении (возвратное движение). Суставы, обеспечивающие более одной степени свободы для звеньев пары, оснащены мышцами, имеющими различные направления тяги. У всех синергистов есть составляющие тяги, направленные в сторону движения. Эти составляющие в сумме создают движущую равнодействующую тягу (D1+D2). Все антагонисты растягиваются при движении. Одна составляющая тяги у них направлена одинаково — против движения (тормозящая тяга — Т] -[ Та). Поскольку антагонисты (как и синергисты) тянут под углом друг к другу, то они имеют не только тормозящие, но и отклоняющие (нормальные) тяги (Рг и /?г)- Изменение соотношения нейтрализующих друг друга (нормальных) составляющих тяги синергистов и антагонистов может изменить направление движения. При этом создается перевес суммарной отклоняющей тяги в ту или иную сторону. Теперь рассмотрим совместное действие всех тяг, приложенных к звену. От соотношения движущей и тормозящей тяг зависят величина равнодействующей (ускоряющей или замедляющей) силы, ускорение звена и, следовательно, величина его скорости, а от соотношения направляющих тяг— величина силы, определяющей направление движения. Изменение направления скорости характерно для многоосных суставов, в которых возможно бесчисленное множество вариантов направления движений. Изменение же величины скорости имеет место во всех суставах — и одноосных, и многоосных. Моментами всех мышечных тяг относительно оси сустава (главным моментом мышц) определяются ускорения звена под действием совокупности работающих мышц. Поскольку моменты тяг изменяются, главный момент мышц непостоянен. Совместная тяга групп мышц и действие других внешних сил для звена сил определяют в любом суставе величину скорости, а в суставах с многими степенями подвижности — еще и направление движения. «Мышцы — главные тормоза движения»,— писал П. Ф. Лесгафт. Именно антагонисты притормаживают движущееся звено тела и останавливают его, совершая упругую амортизацию при перегрузке, предохраняя от разрушения кости, хрящи, связки, суставные сумки. Один из крупнейших русских физиологов А. А. Ухтомский обращал особое внимание на то, что в каждом неодноосном суставе заложены возможности множества механизмов (неполносвязный механизм). Совместное действие групп мышц превращает биокинематическую пару с неодноосным сочленением в биодинамически полно-связный механизм — с единственно возможным направлением движения, т. е. исключением всех остальных. Кроме функциональных групп мышц одного сустава (синергисты, антагонисты) существуют и другие группы совместно действующих мышц. Двусуставные мышцы ряда суставов одной кинематической цепи при движении в одном суставе обусловливают, растягиваясь и напрягаясь, содружественное движение в соседних суставах — это так называемая мышечная координация. Так, при сгибании тазобедренного сустава сгибается коленный (тяги двусуставных мышц задней поверхности бедра) и разгибается голеностопный (тяги мышц передней поверхности голени) суставы. Наконец, в замкнутых цепях можно различать группы совместно действующих мышц, обусловливающих благодаря передаточному действию одновременные движения в суставах замкнутой цепи. 6.3. Взаимодействие групп мышц при разных сопротивлениях Напряжение синергистов при разных сопротивлениях изменяется соответственно изменению сопротивления, антагонисты же напрягаются преимущественно при уменьшающемся сопротивлении (силы инерции), а также при общих больших нагрузках на сустав. Все сопротивления движению по их величине можно разделить на постоянные, увеличивающиеся и уменьшающиеся. Примером постоянных сопротивлений служат вес, сила трения; пример увеличивающихся сопротивлений — нарастающие силы упругой деформации (упражнения с эспандером). В обоих случаях противодействие антагонистов при регулировании величины скорости не является необходимым. Действительно, антагонисты при постоянных и нарастающих сопротивлениях не включаются в работу (пока их сильно не растянут). При уменьшающихся же сопротивлениях (например, силы инерции во время разгона снаряда) антагонисты быстрее вступают в работу — притормаживают звено и этим обеспечивают точность движения, предупреждая травму в случае предельно быстрого окончания движения. Напряжения синергистов в этих трех случаях соответствуют сопротивлениям (постоянные, нарастающие, убывающие—рис. 13, а). Время включения мышц в работу и выключения из нее зависит от конкретных условий движения (см. рис. 13, б). 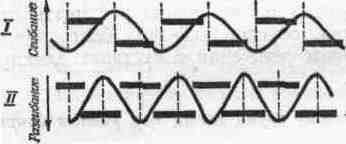 Разгибатель Рис. 13. Взаимодействие антагонистических пар мышц: б — при темпах: I — медленном, // — быстром (по К. Вахгольдеру) Роль инерционных и упругих сил при медленных и быстрых движениях не одинакова. Согласование работы синергистов и антагонистов в зависимости от скорости движения существенно различается. Поэтому участие мышц в медленных и быстрых движениях, даже одинаковых по форме, различно. 6.4. Перераспределение напряжений мышц Моменты включения мышцы в работу и выключения из нее определяются зоной ее активности и оптимальной зоной, что приводит по ходу движения к постоянному изменению тяги мышц — перераспределению напряжений. Каждая мышца имеет в определенном движении свою зону активности, в пределах которой она может выполнять необходимую для этого движения функцию. Границы этой зоны могут составлять пределы возможного размаха движения, или позы, по достижении которых мышца уже не в состоянии осуществлять требуемую для данного движения функцию, хотя звено еще способно продолжать движение в этом направлении. Границы зоны активности в многоосном суставе определяются сменой функций мышц в момент их наименьшей длины. Если движение звена продолжается далее этой границы, то и режим мышцы сменяется (с преодолевающей работы на уступающую, или наоборот). Кроме того, у каждой мышцы есть оптимальная зона действия при определенном режиме работы. В этой зоне ее тяга в данном движении наиболее эффективна. Границы оптимальной зоны зависят от условий эффективности работы мышцы — от того, как она растянута, под каким углом тянет (каково ее плечо силы). В пределах оптимальной зоны различают еще акцентируемый участок,где момент силы тяги мышцы наибольший. Местоположение этого участка зависит часто от согласования движений с движениями других звеньев. При входе в соответствующую зону мышца обычно возбуждается, выходя из нее — выключается из активной деятельности, при этом ее напряжение изменяется. Значит, распределение напряжений в группе мышц определенного сустава по ходу движения изменяетеcя. Нервной системой вносятся необходимые уточнения, как моментов включения мышц в работу и выключения из нее, так и величины напряжения (коррекции напряжения, вызванные координационным несоответствием тяги других мышц сустава). |