кур2. Курсовая работа
 Скачать 411.03 Kb. Скачать 411.03 Kb.
|

|
КУРСОВАЯ РАБОТА Роль фосфолипазы С и сфингомиелиназы в модификации липидного бислоя мембран. Нервные клетки, или нейроны, осуществляют рецепцию электрического сигнала, передачу его другим нервным клеткам или клеткам-эффекторам. Нейроны отличаются большим разнообразием своих размеров, формы, строения, функции и реактивности. Они занимают определенное место в составе рефлекторных дуг, представляющих материальный субстрат рефлексов. В связи с этим по функциональным свойствам различают чувствительные (рецепторные), вставочные (ассоциативные) и двигательные (эффекторные) нейроны. По гистологическим признакам нервные клетки подразделяются на звездчатые, пирамидные, веретеновидные, паукообразные и др. На форму клеток влияют число отростков и способы их отхождения от тела нейрона. Тело нервной клетки содержит нейроплазму и обычно одно ядро. Размер тела варьирует в широких пределах от 5 до 130 мкм. Отростки имеют длину от нескольких микрометров до 1-1,5 м. По количеству отростков выделяют нейроны униполярные (с одним отростком), биполярные (с двумя отростками) и мультиполярные (с числом отростков более двух). Отростки нервных клеток специализированы на выполнение определенных функций и потому подразделяются на два вида. Одни из них называются дендритами, поскольку они сильно ветвятся. Эти отростки воспринимают раздражение и проводят импульсы по направлению к телу нейрона. Отростки другого вида называются аксонами. Они выполняют функцию отведения нервных импульсов от тела нейрона. Нервные клетки имеют несколько дендритов, но один аксон. 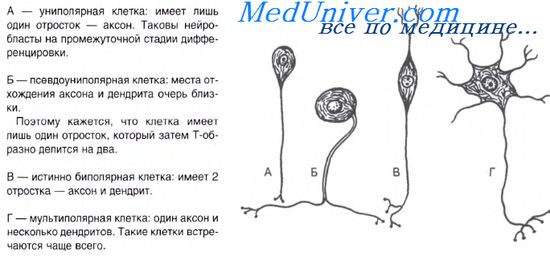 Отростки нервных клеток, покрытые оболочками, называются нервными волокнами. По строению оболочек различают миелиновые и безмиелиновые нервные волокна. Отросток нервной клетки в нервном волокне называют осевым цилиндром, или аксоном, так как чаще всего (за исключением чувствительных нервов) в составе нервных волокон находятся именно аксоны. В центральной нервной системе оболочки отростков нейронов образуются отростками олигодендроглиоцитов, а в периферической — нейролеммоцитами Шванна. Безмиелиновые нервные волокна находятся преимущественно в составе автономной, или вегетативной, нервной системы. Нейролеммоциты оболочек безмиелиновых нервных волокон, располагаясь плотно, образуют тяжи. В нервных волокнах внутренних органов, как правило, в таком тяже имеется не один, а несколько осевых цилиндров, принадлежащих различным нейронам. Они могут, покидая одно волокно, переходить в соседнее. Такие волокна, содержащие несколько осевых цилиндров, называются волокнами кабельного типа. По мере погружения осевых цилиндров в тяж нейролеммоцитов оболочки последних прогибаются, плотно охватывают осевые цилиндры и, смыкаясь над ними, образуют глубокие складки, на дне которых и располагаются отдельные осевые цилиндры. Сближенные в области складки участки оболочки нейролеммоцита образуют сдвоенную мембрану — мезаксон, на которой как бы подвешен осевой цилиндр. Миелиновые нервные волокна встречаются как в центральной, так и в периферической нервной системе. Они значительно толще безмиелиновых нервных волокон. Они также состоят из осевого цилиндра, «одетого» оболочкой из нейролеммоцитов Шванна, но диаметр осевых цилиндров этого типа волокон значительно толще, а оболочка сложнее. Миелиновый слой оболочки такого волокна содержит значительное количество липидов, поэтому при обработке осмиевой кислотой он окрашивается в темно-коричневый цвет. В миелиновом слое периодически встречаются узкие светлые линии—насечки миелина, или насечки Шмидта — Лантермана. Через определенные интервалы (1—2 мм) видны участки волокна, лишенные миелинового слоя, — это так называемые узловатые перехваты, или перехваты Ранвье. Скорость передачи импульса миелиновыми волокнами больше, чем безмиелиновыми. Тонкие волокна, бедные миелином, и безмиелиновые волокна проводят нервный импульс со скоростью 1—2 м/с, тогда как толстые миелиновые — со скоростью 5—120 м/с. В безмиелиновом волокне волна деполяризации мембраны идет по всей аксолемме, не прерываясь, а в миелиновом возникает только в области перехватов. Таким образом, для миелиновых волокон характерно сальтаторное проведение возбуждения, т.е. прыжками. Между перехватами идет электрический ток, скорость которого выше, чем прохождение волны деполяризации по аксолемме. 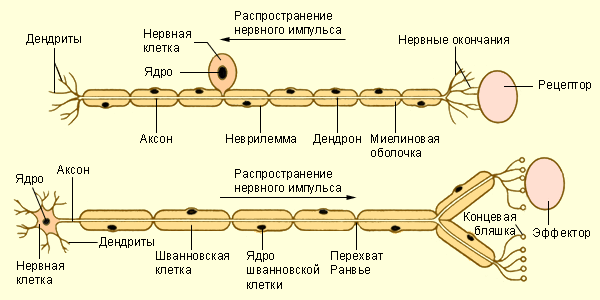 Не́рвные воло́кна — длинные отростки нейронов, покрытые глиальными оболочками. По нервным волокнам распространяются нервные импульсы, по каждому волокну изолированно, не заходя на другие.[1][2]. В различных отделах нервной системы оболочки нервных волокон значительно отличаются по своему строению, что лежит в основе деления всех волокон на миелиновые и безмиелиновые. Те и другие состоят из отростка нервной клетки, лежащего в центре волокна, и поэтому называемого осевым цилиндром (аксоном), и, в случае миелиновых волокон, окружающей его оболочкой. В зависимости от интенсивности функциональной нагрузки нейроны формируют тот или иной тип волокна. Для соматического отдела нервной системы, иннервирующей скелетную мускулатуру, обладающую высокой степенью функциональной нагрузки, характерен миелиновый (мякотный) тип нервных волокон, а для вегетативного отдела, иннервирующего внутренние органы — безмиелиновый (безмякотный) тип. Классификация Эрлангера-Гассера. Является наиболее полной классификацией нервных волокон по скорости проведения нервного импульса.
Классификация по Ллойду. Классифицирует только афферентные нейроны.
Миелинизация нервных волокон. При формировании безмиелинового нервного волокна осевой цилиндр (отросток нейрона) погружается в тяж из леммоцитов, цитолеммы которых прогибаются и плотно охватывают осевой цилиндр в виде муфты, края которой смыкаются над ним, образуя дупликатуру клеточной мембраны — мезаксон. Соседние леммоциты входящие в состав сплошного глиального тяжа своими цитолеммами образуют простые контакты. Безмиелиновые нервные волокна имеют слабую изоляцию, допускающую переход нервного импульса с одного волокна на другое, как в области мезаксона, так и в области межлеммоцитарных контактов. Миелиновые нервные волокна значительно толще безмиелиновых. Принцип образования их оболочек такой же, как и безмиелиновых, то есть осевые цилиндры также прогибают цитолемму глиоцитов, образуя линейный мезаксон. Однако, быстрый рост нейронов соматического отдела нервной системы, связанный с формированием и ростом всего организма, приводит к вытягиванию мезаксонов, многократному обращению леммоцитов вокруг осевых цилиндров. В результате образуются концентрические наслоения. При этом цитоплазма с ядром леммоцитов оттесняется в область последнего витка, образующего наружный слой оболочек волокна, называемой шванновской оболочкой или неврилеммой. Внутренний слой, состоящий из витков мезаксона, называется миелиновым или миелиновой оболочкой. Следствием того, что миелинизация происходит в процессе роста как отростков нейронов, так и самих леммоцитов, является постепенное увеличение количества витков и размеров мезаксона, то есть каждый последующий виток шире предыдущего. Следовательно, последний виток, содержащий цитоплазму с ядром леммоцита является самым широким. Толщина миелина по длине волокна неоднородна, а в местах контактов соседних леммоцитов слоистая структура исчезает и контактируют лишь наружные слои, содержащие цитоплазму и ядро. Места их контактов называются узловыми перехватами (перехватами Ранвье), возникающими вследствие отсутствия здесь миелина и истончения волокна. В ЦНС миелинизация нервного волокна происходит за счет обхвата осевых цилиндров отростками олигодендроцитов. Как мембранная структура миелин имеет липидную основу и при обработке окисями окрашивается в тёмный цвет. Другие компоненты мембраны и промежутки не окрашиваются, поэтому периодически встречаются светлые полоски − насечки миелина (насечки Шмидта-Лантермана), которые соответствуют небольшим прослойкам цитоплазмы леммоцита. В цитоплазме осевого цилиндра располагаются продольно ориентированные нейрофибриллы и митохондрии, которых больше в непосредственной близости к перехватам и в концевых аппаратах волокна. Цитолемма осевого цилиндра (аксона) называется аксолеммой. Она обеспечивает проведение нервного импульса, который представляет собой волну деполяризации аксолеммы. Если осевой цилиндр представлен нейритом, то в нём отсутствуют гранулы базофильного вещества. Мембраны нервных волоконТаким образом, в афферентных нервных волокнах потенциал действия первично возникает на ближайшем к рецептору участке мембраны нервного волокна, имеющем потенциалзависимые натриевые каналы. Возникнув в начале волокна, потенциал действия проводится вдоль него по направлению к телу нейрона и далее к нервным центрам. В сенсорных рецепторах, которые образованы не нервными окончаниями, а целостными нервными или эпителиальными клетками, возникший рецепторный потенциал оказывает возбуждающее действие на чувствительное нервное окончание через синаптическую связь. При возникновении рецепторного потенциала в синаптическую щель выделяется медиатор, который деполяризует постсинаптическую мембрану нервного окончания, и на ближайшем безмиелиновом участке этого волокна возникает потенциал действия, передающийся к нервным центрам. В хеморецепторах механизм генерации рецепторного потенциала несколько отличается от механизма в механорецепторах. Так, в обонятельных рецепторах молекула вещества связывается с чувствительным к нему рецептором, что приводит к активации цепочки биохимических реакций, образующих вещества (так называемые вторичные посредники), которые открывают в мембране рецептора натриевые и кальциевые каналы. Вход в рецепторную обонятельную клетку Na+ и Са2+ обеспечивает генерацию на ее мембране рецепторного потенциала. Особенности строения и виды нервных волоконНервное волокно - аксон - покрыт клеточной мембраной. Выделяют 2 вида нервных волокон. Безмиелиновые нервные волокна - один слой швановских клеток, между ними - щелевидные пространства. Клеточная мембрана на всем протяжении контактирует с окружающей средой. При нанесении раздражения возбуждение возникает в месте действия раздражителя. Безмиелиновые нервные волокна обладают электрогенными свойствами (способностью генерировать нервные импульсы) на всем протяжении. Миелиновые нервные волокна - покрыты слоями шванновских клеток, которые местами образуют перехваты Ранвье (участки без миелина) через каждые 1 мм. Продолжительность перехвата Ранвье 1 мкм. Миелиновая оболочка выполняет трофическую и изолирующую функции (высокое сопротивление). Участки, покрытые миелином не обладают электрогенными свойствами. Ими обладают перехваты Ранвье. Возбуждение возникает в ближайшем к месту действия раздражителя перехвата Ранвье. В перехватах Ранвье высокая плотность Nа-каналов, поэтому в каждом перехвате Ранвье происходит усиление нервных импульсов. Перехваты Ранвье выполняют функцию ретрансляторов (генерируют и усиливают нервные импульсы). Механизм проведения возбуждения по нервному волокну 1885 г. - Л. Герман - между возбужденными и невозбужденными участками нервного волокна возникают круговые токи. При действии раздражителя имеется разность потенциалов между наружной и внутренней поверхностями ткани (участки несущие различные заряды). Между этими участками возникает электрический ток (движение ионов Nа+). Внутри нервного волокна возникает ток от положительного полюса к отрицательному полюсу, т. е. ток направлен от возбужденного участка к невозбужденному. Этот ток выходит через невозбужденный участок и вызывает его перезарядку. На наружной поверхности нервного волокна ток идет от невозбужденного участка к возбужденному. Этот ток не изменяет состояние возбужденного участка, т. к. он находится в состоянии рефрактерности. Доказательство наличия круговых токов: нервное волокно помещают в раствор NaCl и регистрируют скорость проведения возбуждения. Затем нервное волокно помещают в масло (повышается сопротивление) - скорость проведения уменьшается на 30 %. После этого нервное волокно оставляют на воздухе - скорость проведения возбуждения уменьшается на 50 %. Особенности проведения возбуждения по миелиновым и безмиелиновым нервным волокнам:
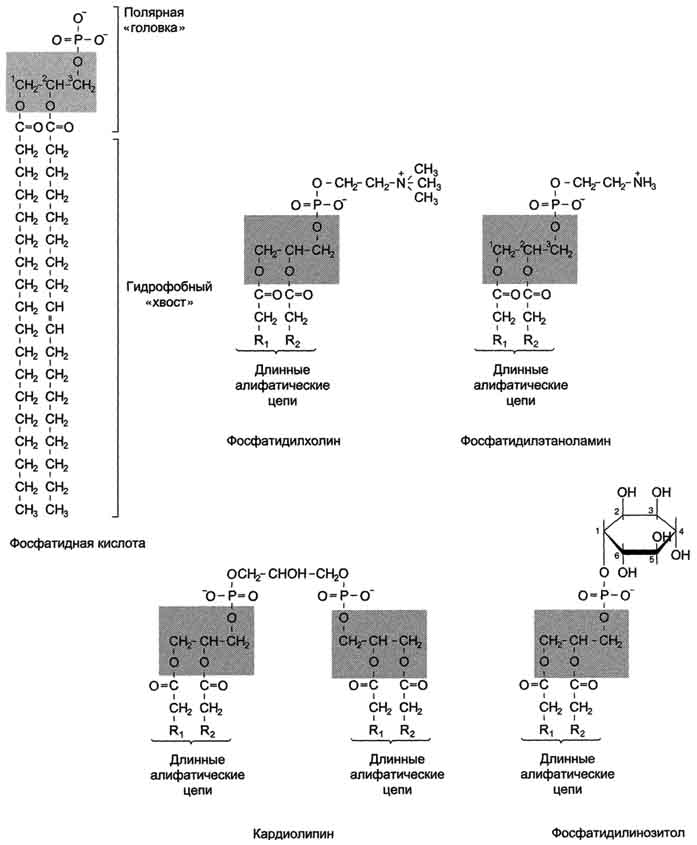
Этот способ менее выгоден: большие затраты энергии (на работу Nа-К-насоса), меньшая скорость проведения возбуждения. 1. Структура и свойства липидов мембран Мембранные липиды - амфифильные (амфипатические) молекулы, т.е. в молекуле есть как гидрофильные группы (полярные "головки"), так и алифатические радикалы (гидрофобные "хвосты"), самопроизвольно формирующие бислой. В большинстве эукариотических клеток они составляют около 30-70% массы мембраны (рис. 5-2). В мембранах присутствуют липиды трёх главных типов - фосфолипиды, гликолипиды и холестерол (холестерин). Липидный состав мембран различен, содержание того или другого липида, по-видимому, определяется разнообразием функций, выполняемых этими липидами в мембранах. Фосфолипиды. Все фосфолипиды можно разделить на 2 группы - глицерофосфолипиды и сфингофосфолипиды. Глицерофосфолипиды относят к производным фосфатидной кислоты. Наиболее распространённые глицерофосфолипиды мембран - фосфатидилхолины и фосфатидилэтаноламины (рис. 5-3). В мембранах эукариотических клеток обнаружено огромное количество разных фосфолипидов, причём они распределены неравномерно по разным клеточным мембранам. Эта неравномерность относится к распределению как полярных "головок" (табл. 5-1), так и ацильных остатков (табл. 5-2). Каждый глицерофосфолипид, например фосфатидилхолин, представлен несколькими десятками фосфатидилхолинов, отличающихся друг от друга строением жирно-кислотных остатков. Рис. 5-2. Содержание липидов и белков в различных клеточных мембранах (%). На долю глицерофосфолипидов (полярная группа - инозитол) приходится лишь 2-8% всех фосфолипидов, содержащихся в клеточной мембране эукариотов. Инозитол в составе фосфати-дилинозитолов может быть фосфорилирован по С4 (фосфатидилинозитол-4-монофосфат) или С4 и С5(фосфатидилинозитол-4,5-бисфосфат). Рис. 5-3. Глицерофосфолипиды мембран.
Таблица 5-1. Фосфолипидный состав клеточных органелл и плазматической мембраны гепатоцитов
Таблица 5-2. Жирно-кислотный состав некоторых мембран печени
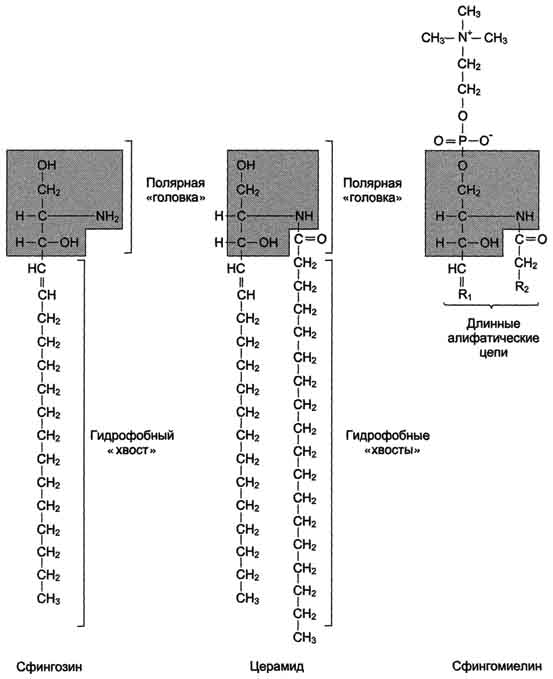
В состав фосфатидш1инозитол-4,5-бисфосфатов входят в основном ацильные остатки стеариновой или пальмитиновой (по первому положению глицерола) и арахидоновой (по второму положению) жирных кислот. Специфические фосфолипиды внутренней мембраны митохондрий - кардиолипины (дифосфатидилглицеролы), построенные на основе глицерола и двух остатков фосфатид-ной кислоты. Они синтезируются ферментами внутренней мембраны митохондрий и составляют около 22% от всех фосфолипидов мембраны. В плазматических мембранах клеток в значительных количествах содержатся сфингомие-лины (рис. 5-4). Сфингомиелины построены на основе церамида - ацилированного аминоспирта сфингозина. Полярная группа состоит из остатка фосфорной кислоты и холина, этаноламина или серина. Сфингомиелины - главные липиды миелиновой оболочки нервных волокон. Гликолипиды. В гликолипидах гидрофобная часть представлена церамидом. Гидрофильная группа - углеводный остаток, присоединённый гликозидной связью к гидроксильной группе у
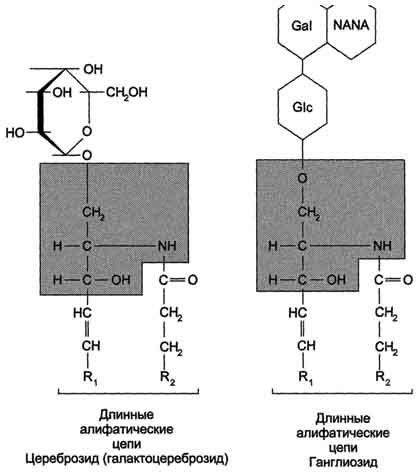
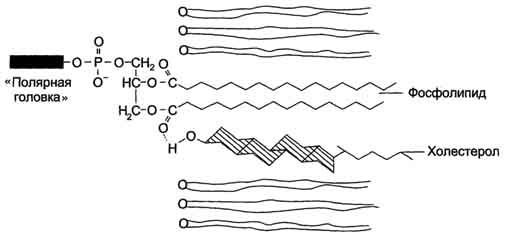
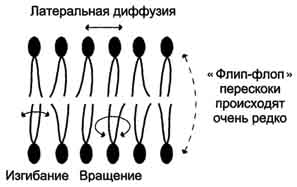
Рис. 5-4. Сфингофосфолипиды - производные церамида. первого углеродного атома церамида (рис. 5-5). В зависимости от длины и строения углеводной части различают цереброзиды, содержащие моно- или олигосахаридный остаток, и ганглиозиды, к ОН-группе которых присоединён сложный, разветвлённый олигосахарид, содержащий N-ацетилнейраминовую кислоту (NANA) (см. раздел 7). Полярные "головки" гликосфинголипидов находятся на наружной поверхности плазматических мембран. В значительных количествах гликолипиды содержатся в мембранах клеток мозга, эритроцитов, эпителиальных клеток. Ганглиозиды эритроцитов разных индивидуумов различаются строением олигосахаридных цепей, проявляющих антигенные свойства. Рис. 5-5. Гликолипиды. Gal - галактоза; Glc - глюкоза; NANA (NeuAc) - N-ацетилнейраминовая или сиаловая кислота. Холестерол. Холестерол присутствует во всех мембранах животных клеток. Его молекула состоит из жёсткого гидрофобного ядра и гибкой углеводородной цепи, единственная гидроксильная группа является "полярной головкой" (рис. 5-6). Для животной клетки среднее молярное отношение холестерол/фосфолипиды равно 0,3-0,4, но в плазматической мембране это соотношение гораздо выше (0,8-0,9). Наличие холестерола в мембранах уменьшает подвижность жирных кислот, снижает латеральную диффузию липи-дов и белков, и поэтому может влиять на функции: мембранных белков. В составе мембран растений холестерола нет, а присутствуют растительные стероиды - ситостерол и стигмастерол. 2. Трансмембранная асимметрия липидов Каждая мембрана клетки замкнута, т.е. имеет внутреннюю и внешнюю поверхности, различающиеся по липидному и белковому составам - эту особенность мембран называюттрансмембранной (поперечной) асимметрией. Липидная асимметрия возникает прежде всего потому, что липиды с более объёмными полярными "головками" стремятся находиться в наружном монослое, так как там площадь поверхности, приходящаяся на полярную "головку", больше. Фосфатадилхолины и сфингомиелины локализованы преимущественно в наружном монослое, а фосфатидилэтаноламины и фосфатидилсерины в основном во внутреннем. Липиды в некоторых биологических мембранах с довольно большой частотой мигрируют с одной стороны мембраны на другую, т.е. совершают "флип-флоп" (от англ, flip-flop) перескоки (рис. 5-7). Перемещение липидных молекул затрудняют полярные "головки", поэтому липиды, находящиеся на внутренней стороне мембраны, имеют относительно высокую скорость Рис. 5-6. Положение молекулы холестерола в мембране. Молекула холестерола располагается в липидном слое мембраны параллельно алифатическим цепям молекул фосфо- и гликолипидов. Гидроксильная группа холестерола контактирует с гидрофильными "головками" этих липидов. Рис. 5-7. Типы движений липидных молекул в бислое мембран. трансмембранной миграции по сравнению с липидами наружной стороны мембраны, мигрирующих медленнее или вообще не совершающими "флип-флоп" перескоки. 3. Жидкостностъ мембран Для мембран характерна жидкостность (текучесть), способность липидов и белков к латеральной диффузии. Скорость перемещения молекул зависит от микровязкости мембран, которая, в свою очередь, определяется относительным содержанием насыщенных и ненасыщенных жирных кислот в составе липидов. Микровязкость меньше, если в составе липидов преобладают ненасыщенные жирные кислоты, и больше при высоком содержании насыщенных жирных кислот. Ацильные (алифатические) остатки ненасыщенных жирных кислот имеют так называемые "изломы" (см. раздел 8). Эти "изломы" препятствуют слишком плотной упаковке молекул в мембране и делают её более рыхлой, а следовательно и более "текучей". На текучесть мембран также влияют размеры углеводородных "хвостов" липидов, с увеличением длины которых мембрана становится более "текучей". 4. Функции мембранных липидов Фосфо- и гликолипиды мембран, помимо участия в формировании липидного бислоя, выполняют ряд других важных функций. Липиды формируют среду для функционирования Мембранных белков, принимающих в ней нативную конформацию. Выделенные из мембран ферменты, лишённые липидного окружения, как правило, не проявляют каталитической активности. Некоторые мембранные липиды - предшественники вторичных посредников при передаче гормонального сигнала. Так, фосфати-дилинозитол-4,5-бисфосфат (ФИФ2) под действием фермента фосфолипазы С гидролизуется до диацилглицерола (ДАТ), активатора протеинкиназы С и инозитол-1,4,5-трифосфата (ИФ3) - регулятора кальциевого обмена в клетке (рис. 5-8). ДАГ, ИФ3, протеинкиназа С и Са2+ - участники инозитолфосфатной системы передачи сигнала. Кроме того, некоторые липиды выполняют "якорную" функцию, например к фосфатидил-инозитолам через олигосахарид могут присоединяться специфические белки наружной поверхности
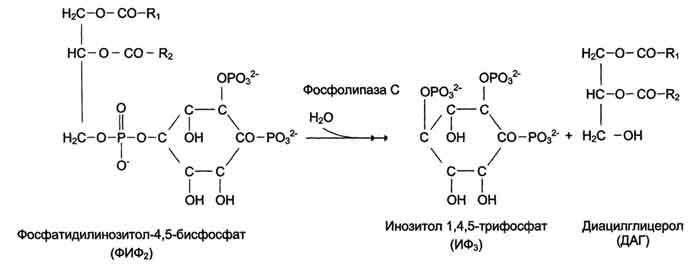 Рис. 5-8. Гидролиз фосфатидилинозитол-4,5-бисфосфата. клетки (рис. 5-9). Фосфатидилинозитол с присоединённым к нему олигосахаридом (гликаном) называют фосфатидилинозитолгликаном. Связь белков с этой молекулой (гликаном) осуществляется через фосфоэтаноламин. Пример такого "заякоренного" белка - ацетилхолинэстераза, катализирующая гидролиз ацетилхолина в синаптической щели. Этот фермент фиксируется на поСтеинаптической мембране, ковалентно присоединяясь к фосфатидилино-зитолгликану. Под действием фосфолипазы С может происходить отделение белков от внешней поверхности клетки. Липиды могут быть аллостерическими активаторами мембранных ферментов. Например, р-гидроксибутиратдегидрогеназа, участвующая в окислении кетоновых тел (см. раздел 8), локализована на внутренней мембране митохондрий. Каталитическая активность фермента проявляется только в присутствии фосфатидилхолина. Фермент протеинкиназа С катализирует реакции фосфорилирования белков по аминокислотным 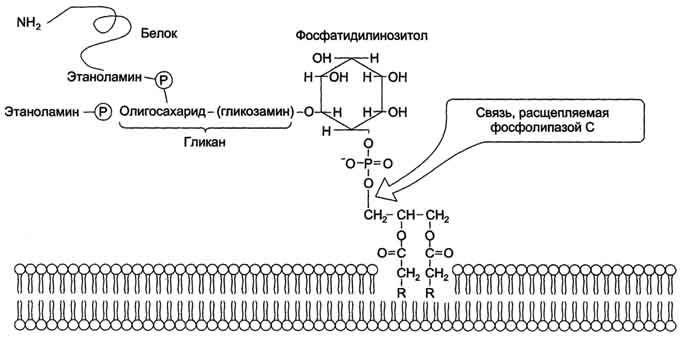 Рис. 5-9. "Якорная" функция фосфатидилинозитолгликанов.
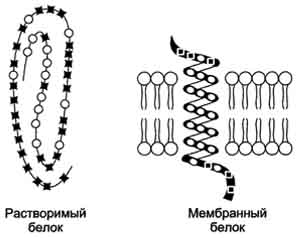
остаткам серина и треонина. В неактивной форме протеинкиназа С находится в цитозоле. Однако после стимуляции клетки (повышение в клетке концентрации кальция) фермент быстро активируется ионами кальция и оказывается связанным с мембраной. Функционально активная протеинкиназа С - комплекс, содержащий мономер фермента, молекулу диацилглицерола, один или более ионов Са2+ и четыре молекулы фосфатидилсерина. Креатинкиназа, фермент катализирующий образование макроэргического соединения креа-тинфосфата (см. раздел 9). Для проявления его активности требуется специфическое взаимодействие с кардиолипином внутренней мембраны митохондрий. БЕЛКИ МЕМБРАН Если основная роль липидов в составе мембран заключается в стабилизации бислоя, то белки отвечают за функциональную активность мембран. Одни из них обеспечивают транспорт определённых молекул и ионов, другие являются ферментами, третьи участвуют в связывании цитоскелета с внеклеточным матриксом или служат рецепторами для гормонов, медиаторов, эйкозаноидов, липопротеинов, оксида азота (NO). На долю белков приходится от 30 до 70% массы мембран. Белки определяют особенности функционирования каждой мембраны. Особенности строения и локализации белков в мембранах Мембранные белки, контактирующие с гидрофобной частью липидного бислоя, должны быть амфифильными. Те участки белка, которые взаимодействуют с углеводородными цепями жирных кислот, содержат преимущественно неполярные аминокислоты. Участки белка, находящиеся в области полярных "головок", обогащены гидрофильными аминокислотными остатками. Белки мембран различаются по своему положению в мембране (рис. 5-10). Они могут глубоко проникать в липидный бислой или даже пронизывать его - интегральные белки, либо разными способами прикрепляться к мембране - поверхностные белки. Поверхностные белки Поверхностные белки часто прикрепляются к мембране, взаимодействуя с интегральными 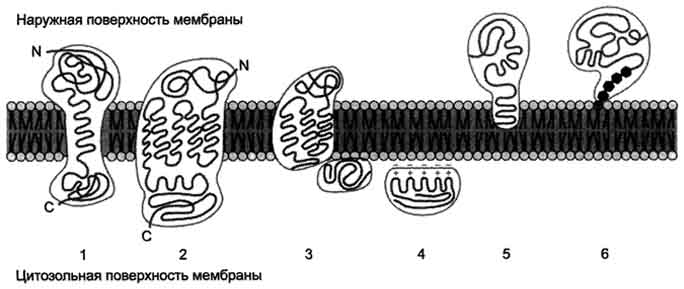 Рис. 5-10. Расположение (локализация) белков в мембранах. Трансмембранные белки, например: 1 - гпикофорин А; 2 - рецептор адреналина. Поверхностные белки: 3 - белки, связанные с интегральными белками, например, фермент сукци-натдегидрогеназа; 4 - белки, присоединённые к полярным "головкам" липидного слоя, например, протеинкиназа С; 5 - белки, "заякоренные" в мембране с помощью короткого гидрофобного концевого домена, например, цитохром b5; 6 - "заякоренные" белки, ковалентно соединённые с липидом мембраны (например, фермент щелочная фосфатаза).
белками или поверхностными участками липидного слоя. Белки, образующие комплексы с интегральными белками мембраны Ряд пищеварительных ферментов, участвующих в гидролизе крахмала и белков, прикрепляется к интегральным белкам мембран микроворсинок кишечника. Примерами таких комплексов могут быть са-хараза-изомальтаза и мальтаза-гликоамилаза (см. раздел 7). Возможно, связь этих пищеварительных ферментов с мембраной позволяет с высокой скоростью гидролизовать субстраты и усваивать продукты гидролиза клеткой. Белки, связанные с полярными «головками» липидов мембран Полярные или заряженные домены белковой молекулы могут взаимодействовать с полярными "головками" липидов, образуя ионные и водородные связи. Кроме того, множество растворимых в цитозоле белков при определённых условиях могут связываться с поверхностью мембраны на непродолжительное время. Иногда связывание белка -- необходимое условие проявления ферментативной активности. К таким белкам, например, относят протеинкиназу С, факторы свёртывания крови. Закрепление с помощью мембранного "якоря" "Якорем" может быть неполярный домен белка, построенный из аминокислот с гидрофобными радикалами. Примером такого белка может служить цитохром b5 мембраны ЭР. Этот белок участвует в окислительно-восстановительных реакциях, как переносчик электронов (см. раздел 12). Роль мембранного "якоря" может выполнять также ковалентно связанный с белком остаток жирной кислоты (миристиновой - С14 или пальмитиновой - С16). Белки, связанные с жирными кислотами, локализованы в основном на внутренней поверхности плазматической мембраны. Миристиновая кислота присоединяется к N-концевому глицину с образованием амидной связи. Пальмитиновая кислота образует тиоэфирную связь с цистеином или сложноэфирную с остатками серина и треонина. Небольшая группа белков может взаимодействовать с наружной поверхностью клетки с помощью ковалентно присоединённого к С-концу белка фосфатидилинозитолгликана. Этот "якорь" - часто единственное связующее звено между белком и мембраной, поэтому при действии фосфолипазы С этот белок отделяется от мембраны. Трансмембранные (интегральные) белки Некоторые из трансмембранных белков пронизывают мембрану один раз (гликофорин), другие имеют несколько участков (доменов), последовательно пересекающих бислой (рис. 5-11). Трансмембранные домены, пронизывающие бислой, имеют конформацию α-спирали. Полярные остатки аминокислот обращены внутрь глобулы, 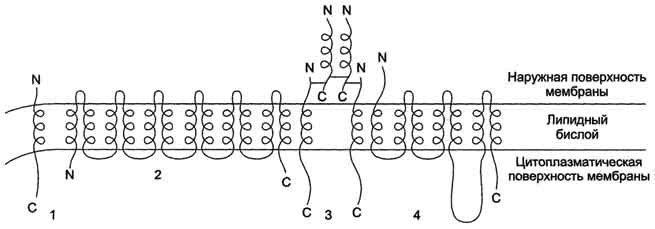 Рис. 5-11. Интегральные белки мембран, содержащие от 1 до 12 трансмембранных доменов. 1- рецептор ЛПНП; 2 - ГЛЮТ-1 - транспортёр глюкозы; 3 - рецептор инсулина; 4 - адренорецептор. а неполярные контактируют с мембранными липидами. Такие белки называют "вывернутыми" по сравнению с растворимыми в воде белками, в которых большинство гидрофобных остатков аминокислот спрятано внутрь, а гидрофильные располагаются на поверхности (рис. 5-12). Радикалы заряженных аминокислот в составе этих доменов лишены заряда и протонированы (-СООН) или депротонированы (-NH2). Гликозилированные белки Поверхностные белки или домены интегральных белков, расположенные на наружной поверхности всех мембран, почти всегда гликозилированы. Олигосахаридные остатки могут быть присоединены через амидную группу аспарагина или гидроксильные группы серина и треонина (рис. 5-13). Олигосахаридные остатки защищают белок от протеолиза, участвуют в узнавании лигандов или адгезии. Латеральная диффузия белков Некоторые мембранные белки перемещаются вдоль бислоя (латеральная диффузия) или поворачиваются вокруг оси, перпендикулярно его поверхности. Например, фермент фосфолипаза А2, связываясь с цитоплазматической поверхностью мембраны, может латерально перемещаться по поверхности бислоя и гидролизовать несколько Рис. 5-12. Локализация неполярных (незакрашенные кружки) и полярных (закрашенные квадраты) аминокислот в растворимых и мембранных белках. тысяч фосфолипидов в минуту до тех пор, пока не отделится от мембраны. Латеральная диффузия интегральных белков в мембране ограничена, это связано с их большими размерами, взаимодействием с другими мембранными белками, элементами цитоскелета или внеклеточного матрикса. Белки мембран не совершают перемещений с одной стороны мембраны на другую ("флип-флоп" перескоки), подобно фосфолипидам. Библиография: 1. Л.В. Высоцкая, Г.М.Дымщиц, Е.М.Низовцев. Общая биология. - М.: Научный мир, 2001.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||