Лекция 1-4. Лекция 1 Физиология, ее предмет, роль в системе медицинского образования и задачи
 Скачать 0.84 Mb. Скачать 0.84 Mb.
|

Изменение МП при действии подпороговых раздражителейПри действии подпороговых раздражителей не происходит возбуждения, но это не значит, что возбудимая ткань не реагирует на действие раздражителя. Наблюдаются следующие явления: электротонический потенциал и локальный ответ. Они развиваются в результате действия раздражителей разной величины и их действие зависит от порога деполяризации. Порог деполяризации – это величина, на которую нужно уменьшить мембранный потенциал, для того чтобы возник потенциал действия. Электротонический потенциалПри действии слабых раздражителей, величина которых не превышает 50% пороговой величины, наблюдается пассивная электротоническая деполяризация или пассивный электротонический потенциал. При этом деполяризация мембраны отмечается только во время действия раздражителя. Развитие и исчезновение электротонического потенциала происходит по прямолинейной (экспоненциальной) кривой. Локальный ответПри увеличении силы подпороговых раздражений от 50 до 99% порога можно наблюдать, что развитие деполяризации происходит не прямолинейно, а по S-образной кривой. Деполяризация продолжает нарастать после прекращения раздражения, а затем сравнительно медленно исчезает. Этот процесс получил название локального ответа (рис.8). Локальный ответ имеет следующие свойства:
Во время развития локального ответа изменяется ионная проницаемость мембраны, увеличивается поток ионов натрия из межклеточной среды в цитоплазму и повышается возбудимость клетки. Изменение возбудимости клетки во время ее возбуждения При возбуждении свойства возбудимой клетки быстро и сильно меняются. Впервые это было замечено Ферворном, который в ответ на действие повторного раздражителя не наблюдал возбуждения. При дальнейшем изучении им была установлена зависимость возбудимости ткани и представлена в виде кривой, которая впоследствии названа кривой возбудимости Ферворна (рис. 10). Допустим, что возбудимость ткани в покое равна 100%. Действуем на нерв или мышцу 2-мя следующими друг за другом раздражителями. В ответ на первый стимул мышца сокращается, а на второй вообще не реагирует, как бы не увеличивать его силу. Это связано с тем, что во время возбуждения возбудимость ткани падает до 0%. Время, в течение которого ткань не реагирует на повторные раздражители, называют абсолютной рефрактерностью. Эта фаза очень короткая: в нервных волокнах она равна 0,001 сек., в скелетных мышцах - 0,005 сек, в сердце - 0,27 сек., самая продолжительная. После этой фазы возбудимость постепенно начинает восстанавливаться, постепенно приближаясь к 100%. Этот период называют фазой относительной рефрактерности. Согласно этому кривая имеет несколько фаз: 1) Кратковременное повышение возбудимости развивается сразу поле действия стимула и совпадает с локальным ответом вначале деполяризации клеточной мембраны. Она связана с постепенным повышением проницаемости натриевых каналов. 1) фаза абсолютной рефрактерности – это полная невозбудимость. Возбудимость клетки резко снижается до 0 . Совпадает с фазой деполяризации на кривой ПД и возникает вследствие полного открытия натриевых каналов, после чего увеличить натриевый ток невозможно, а, значит, клетка не будет отвечать на действие даже сверхпороговых раздражителей. 2) фаза относительной рефрактерности - это период восстановления возбудимости клетки. В эту фазу действие надпороговых раздражителей может вызвать возникновение возбуждения. Она соответствует быстрой реполяризации на кривой ПД и возникает за счет постепенного восстановления натриевых каналов при возвращении заряда клетки к исходной поляризации. 3) фаза экзальтации – возбудимость повышена на 10-20%. Она характеризуется тем, что ткань возбуждается при действии подпороговых стимулов. Эта фаза совпадает со следовой деполяризацией, когда свойства мембраны восстановились, а заряд мембраны еще не достиг своего первоначального уровня. Вследствие этого клетка деполяризована и мембранный потенциал ближе к Екр, поэтому действие подпороговых раздражителей вызовет ПД. 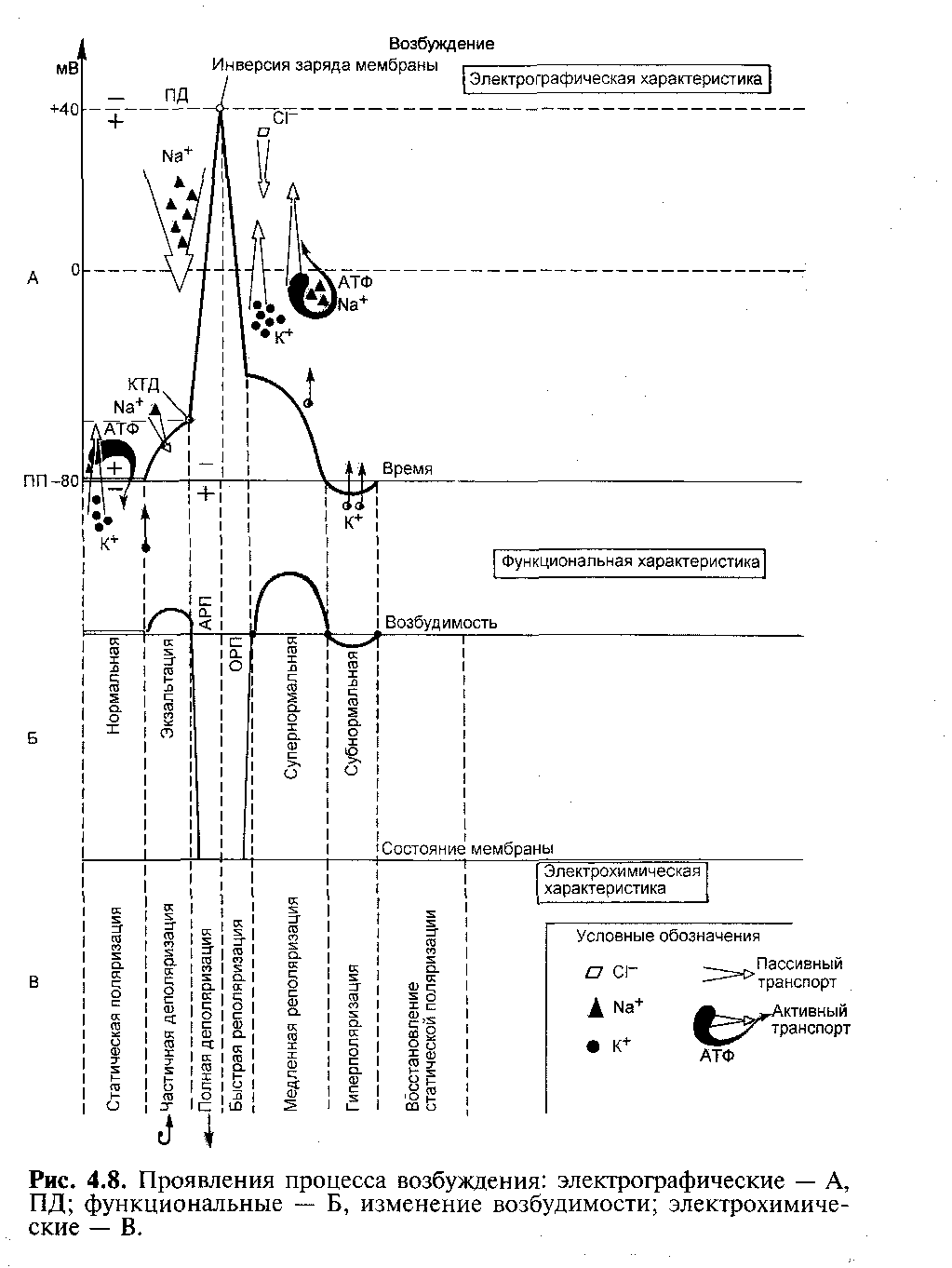 Рис.10 Проявление процесса возбуждения: А - электрографические, ПД; Б - функциональные, изменение возбудимости; В - электрохимические (К.В. Судаков) 4) фаза субнормальной возбудимости – это снижение возбудимости клетки на 10-20%, совпадающее с фазой следовой гиперполяризации. Она связана с избыточным выходом ионов натрия из клетки и с увеличением порога деполяризации, так как мембранный потенциал больше и, соответственно дальше отстоит от Екр. Функциональная лабильность. Максимальный и оптимальный ритмы. Усвоение ритма Н.Е. Введенский (1886) на нервно-мышечном препарате установил, нерв, мионевральные синапсы и мышца по-разному реагируют на сверхпороговые раздражения различной частоты. Так, оказалось, что нерв способен воспроизводить наиболее широкий диапазон частотных раздражений, средний воспроизводится мышцей и наименьший - синапсами. Отсюда был сделан вывод, что разные структуры имеют неодинаковую функциональную подвижность или лабильность (от лат. labilis - скользящий, неустойчивый). Лабильность - это скорость протекания процессов возбуждения в возбудимых тканях. Н.Е Введенский считал мерой лабильности максимальную частоту циклов возбуждения, воспроизводимых в единицу времени (секунду). Таким образом, зная величину лабильности, можно определить полную длительность одного цикла возбуждения. Лабильность находится в прямой зависимости от фазы абсолютной рефрактерности: чем она короче, тем больше лабильность и наоборот. Поэтому лабильность любой возбудимой ткани можно высчитать, зная величину абсолютной рефрактерности. Так, в нервах она длится 0,001 сек, а отсюда огромная лабильность нерва - 1000 имп/сек. В скелетных мышцах рефрактерная фаза составляет 0,004-0,005 сек, а лабильность - 250-300 имп/сек. Очень низка лабильность сердца, всего 3 имп/сек, так как здесь очень продолжительная рефрактерная фаза (0,3 сек). Следовательно, фаза абсолютной рефрактерности ограничивает лабильность ткани. Однако, доказано, что при частоте раздражения с интервалом абсолютной рефрактерности возникает лишь 1 ПД, ткань реагирует лишь на первый стимул, а на последующие - не отвечает. Это связано с тем, что сверхчастые раздражители удлиняют рефрактерность в ткани и развивается катодическая депрессия (пессимум) и восстановление мембранного потенциала после первого ПД не происходит. Поэтому, для получения максимальной частоты ритмические раздражители можно наносить с интервалом, превышающим рефрактерную фазу приблизительно в 2 раза. Поэтому, максимальная частота возбуждений в нервах составляет не 1000, а 500 имп/сек; в мышцах 100-125 имп/сек. Отсюда ясно, что максимальная частота возбуждения представляет крайнюю форму активности и может воспроизводится лишь в особых условиях и весьма короткое время. Следовательно, максимальным называется ритм, который генерирует ткань в экстремальных условиях и короткое время. Даже возбуждение с частотой в 2 фазы абсолютной рефрактерности оказывается слабым, так как он формируется в фазу относительной рефрактерности, когда возбудимость полностью еще не восстановлена. Л.В. латманизова, ученица Ухтомского, обнаружила, что в естественных условиях возбудимые системы функционируют с намного меньшей частотой, чем максимальная. Но эта частота воспроизводится длительно и стойко, без утомления. Тот ритм возбуждения, в котором ткань функционирует длительное время и без утомления, называется оптимальным ритмом. Возникновение оптимального ритма связано с тем, что последующий раздражитель попадает в фазу супернормальной возбудимости, экзальтации, что благоприятствует возникновению возбуждения, и при этом может возникать максимальное сокращение мышцы. Частота, которая вызывает максимальный сократительный эффект, была названа Введенским оптимальным ритмом раздражения, а сокращение - оптимальным. Этот ритм обычно наблюдается и в естественных условиях, возникая в период повышенной возбудимости ткани. Максимальный и оптимальный ритмы связаны между собой математической зависимостью. Оптимальный ритм приблизительно в 5-10 раз реже, чем максимальный. Так для нерва максимальная частота составляет 1000 имп/сек, а оптимальная - 50-100 имп/сек. Для скелетных мышц они соответственно равны 250 и 50 имп/сек. Таким образом, различают две частотные характеристики тканей. Максимальный ритм является мерой лабильности и проявляется в экстренных условиях короткое время. Вторая частотная характеристика - оптимальный ритм используется тканями в естественных условиях. Максимальным ритмом пользуются для выяснения и сравнения лабильности, а оптимальным - для характеристики изменений в функции данной возбудимой системы. В процессе филогенеза лабильность тканей увеличилась. Лабильность ЦНС позвоночных несравненно выше, чем у беспозвоночных. Наиболее она высока в нервах, обеспечивающих срочную связь в организме. В эволюции физиологии есть яркие примеры изменения лабильности определенных органов в зависимости от их роли в жизни животного. Так, у млекопитающих ресничная (цилиарная) мышца является гладкомышечной структурой, поэтому изменение формы хрусталика и аккомодация глаза осуществляется очень медленно. У хищных птиц эта мышца - поперечно-полосатая и с высокой быстротой меняет форму хрусталика, обеспечивая ясное видение при падении птиц на добычу. Лабильность меняется в процессе онтогенеза: при рождении она мала, в 20-30 лет достигает максимума, а после 60 лет быстро снижается, что соответствующим образом меняет работоспособность. Лабильность неодинакова не только у разных тканей, но и у разных структурных единиц одной и той же ткани. Более того, даже у клетки лабильность непостоянна и определяется ее функциональным состоянием. Она может изменяться в процессе длительного воздействия раздражителей. Это, в частности, подтверждается способностью ткани повышать свою функциональную подвижность в процессе жизнедеятельности. При этом у ткани возникают новые свойства, и она приобретает способность воспроизводить более высокий ритм раздражения. Это явление, наблюдаемое в тканях, исследовал ученик и последователь введенского, академик А.А. ухтомский, и назвал процессом усвоения ритма. Повышением лабильности объясняется втягивание в работу, усвоение все большего ритма, а в результате укорочения рефрактерной фазы она способна возрасти в 2 раза. Это связано с ускорением процессов выкачивания ионов натрия из цитоплазмы и более быстрым восстановлением мембранного потенциала. Биопотенциалы гландулоцитов К возбудимым тканям кроме мышечной и нервной тканей относятся и клетки железистого эпителия – гландулоциты, специфическим ответом ко- второго является секреция. Весь секреторный цикл занимает 40-90 минут. Мембранный потенциал различных секреторных клеток равен от –30 до – 75 мВ. Действие раздражителя на гландулоцит меняет мембранный потенциал. Это изменение поляризации клетки называется секреторным потенциалом. Возбуждение в секреторных клетках может иметь два механизма:
В основе деполяризации мембраны лежит повышение проницаемости мембраны для ионов натрия и калия (натрий поступает в железистую клетку, а калий покидает ее). Гиперполяризация мембран происходит за счет поступления ионов хлора в клетку и выходом из нее ионов калия и натрия. Потенциал секреторной клетки сопровождается повышением осмотического давления цитоплазмы и транспортом воды в клетку. Базальная и апикальная мембраны имеют различный заряд в состоянии покоя. Разница в поляризации между апикальной и базальной мембраной в 2-3 мВ создает значительное электрическое напряжение. При возбуждении секреторной клетки оно возрастает и это способствует перемещению секреторных гранул к апикальному полюсу клетки и выходу его наружу. |