Лекция 1-4. Лекция 1 Физиология, ее предмет, роль в системе медицинского образования и задачи
 Скачать 0.84 Mb. Скачать 0.84 Mb.
|

Лекция 3.Физиология скелетных мышцМышечные волокна Скелетные мышцы – это сложное образование, структурной единицей которого являются мышечные волокна, диаметр которых составляет 10-100 мкм. Длина мышечных волокон различна и варьирует от 5до 400 мм. Отдельные мышечные волокна могут быть почти такой же длины, как и мышца. Но чаще всего они короче, так как имеют косое расположение, например, в перистых мышцах. Мышечные волокна объединены в пучки, обвитые соединительной тканью. Внутри пучков находятся клетки, принадлежащие различных двигательным единицам. Двигательная единица – это структура включающая в себя -мотонейрон и иннервируемые им мышечные волокна. У человека количество волокон в мышце устанавливается через 4-5 месяцев после рождения и затем практически не меняется на протяжении всей жизни. При рождении толщина их составляет примерно 1/5 толщины волокон у взрослых людей. Диаметр мышечных волокон может значительно меняться под воздействием тренировки. Нервно-мышечный синапсМ 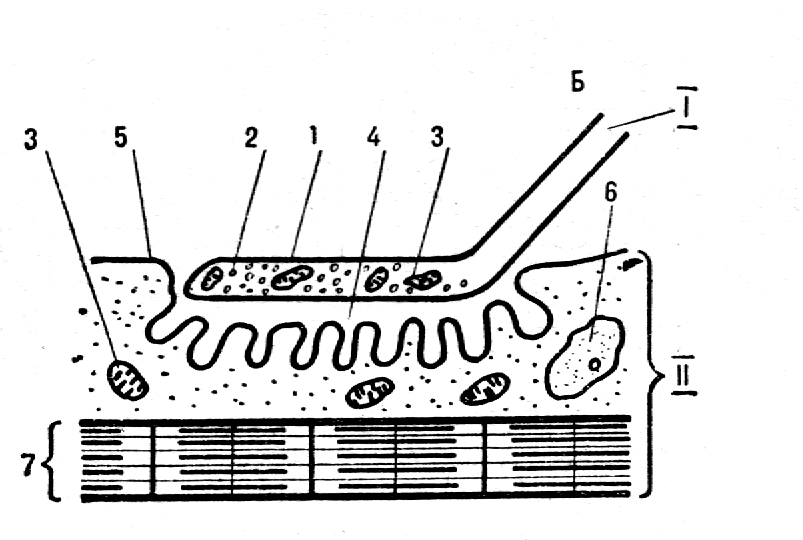 отонейрон связан с мышечным волокном при помощи нервно-мышечного синапса (рис.15). Он имеет две основные части – нервную (пресинаптическую) и мышечную (постсинаптическую). Пресинаптическая часть представлена концевой веточкой аксона, погруженного в углубление на поверхности мышечного отонейрон связан с мышечным волокном при помощи нервно-мышечного синапса (рис.15). Он имеет две основные части – нервную (пресинаптическую) и мышечную (постсинаптическую). Пресинаптическая часть представлена концевой веточкой аксона, погруженного в углубление на поверхности мышечного волокна. Поверхностная мембрана концевой веточки носит название пресинаптическая мембрана. Нервное окончание содержит более миллиона пузырьков ацетилхолина (АХ) – медиатора нервно-мышечного синапса. Мембрана, которая покрывает мышечное волокно в области синапса, называется постсинаптическая мембрана или концевая пластинка. Она образует многочисленные складки, уходящие вглубь волокна, благодаря чему увеличивается ее поверхность. Постсинаптическая мембрана имеет холинорецепторные (хемозависимые) участки и содержит фермент ацетилхолинэстеразу. Между пресинапсом и постсинапсом располагается синаптическая щель. Располагающиеся вокруг синапса шванновские клетки хорошо закрывают синаптическую щель со всех сторон и вещества из межклеточного пространства не могут оказывать влияние на процессы, происходящие в синапсе. Моторная пластинка определяет характеристики двигательной единицы (ДЕ) и имеет следующие особенности:
Мембрана мышечного волокна в состоянии покоя поляризована. Средняя величина потенциала покоя равна 87,2 мВ. Возникший в -мотонейроне нервный импульс распространяется до пресинаптического окончания. Это приводит к тому, что в пресинапсе изменяется проницаемость для ионов натрия и кальция. В результате в синаптическую щель из везикул выделяется медиатор ацетилхолин, который взаимодействует с холинорецепторами хемотропной части мембраны. Он повышает проницаемость для ионов натрия, вызывая деполяризацию постсинаптической мембраны - потенциал концевой пластинки (ПКП). ОН может достигать 60 мВ. М 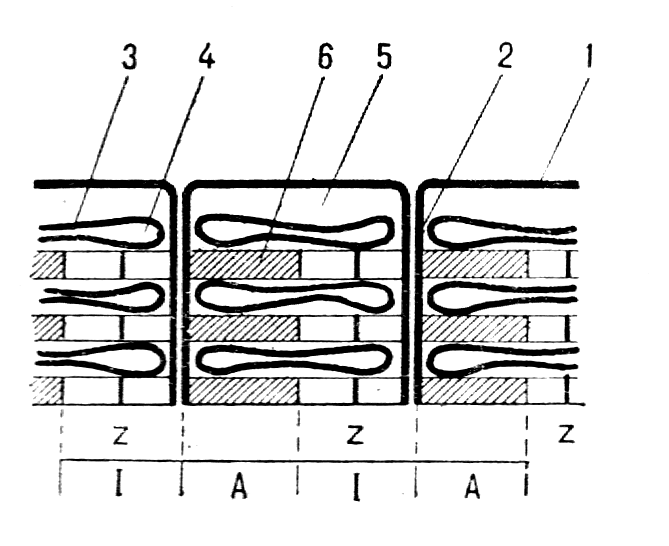 ежду хемозависимой и электрозависимой частями мембраны мышечного волокна развиваются локальные токи, которые, перемещаясь от + к – в обе стороны, вызывают возбуждение всего мышечного волокна. Скорость распространения токов равна в среднем 3,7 м/с. ежду хемозависимой и электрозависимой частями мембраны мышечного волокна развиваются локальные токи, которые, перемещаясь от + к – в обе стороны, вызывают возбуждение всего мышечного волокна. Скорость распространения токов равна в среднем 3,7 м/с.Сокращение мышечных волокон начинается с места контакта аксона и мышцы. Оно также распространяется в оба конца мышечного волокна. В каждом участке волокна проходит определенное латентное время от момента деполяризации до момента наступления сокращения. Латентное время связано с процессом передачи возбуждения от мембраны к сократительным белкам мышечных волокон. Передача происходит с участием саркоплазматического ретикулума (СПР), который состоит из трубочек, начинающихся от мембраны волокна (рис.16). Они расположены поперек мышечного волокна и обвивают каждую миофибриллу в месте соединения А- и I-дисков каждого саркомера. Саркоплазматический ретикулум имеет множество пузырьков, которые окружают каждую миофибриллу по всей длине. Пузырьки контактируют с поперечными канальцами и вместе с ними составляют триады. Потенциал действия передается не только по внешней мембране мышечных волокон, но и по саркоплазматическому ретикулуму к миофибриллам. Ионные механизмы деполяризации саркоплазматического ретикулума отличаются от деполяризации мембраны волокна. Для деполяризации поверхностной мембраны главными являются ионы натрия, тогда как для деполяризации ретикулума – ионы кальция и магния. В состоянии покоя благодаря кальциевому насосу ионы кальция накоплены в саркоплазматическом ретикулуме. Классификация скелетных мышечных волокон Скелетная мускулатура человека и позвоночных животных состоит из мышечных волокон нескольких типов, отличающихся друг от друга структурно-функциональными характеристиками. В настоящее время выделяют четыре основных типа мышечных волокон. Медленные фазические волокна окислительного типа. Волокна этого типа характеризуются большим содержанием белка миоглобина, который способен связывать О2 (близок по своим свойствам к гемоглобину). Мышцы, которые преимущественно состоят из волокон этого типа, за их темно-красный цвет называют красными. Они выполняют очень важную функцию поддержания позы человека и животных. Предельное утомление у волокон данного типа и, следовательно, мышц наступает очень медленно, что обусловлено наличием миоглобина и большого числа митохондрий. Восстановление функции после утомления происходит быстро. Нейромоторные единицы этих мышц состоят из большого числа мышечных волокон. Быстрые фазические волокна окислительного типа. Мышцы, которые преимущественно состоят из волокон этого типа, выполняют быстрые сокращения без заметного утомления, что объясняется большим количеством митохондрий в этих волокнах и способностью образовывать АТФ путем окислительного фосфорилирования. Как правило, число волокон, входящих в состав нейромоторной единицы, в этих мышцах меньше, чем в предыдущей группе. Основное назначение мышечных волокон данного типа заключается в выполнении быстрых, энергичных движений. Быстрые фазические волокна с гликолитическим типом окисления. Волокна данного типа характеризуются тем, что АТФ в них образуется за счет гликолиза. Волокна этой группы содержат митохондрий меньше, чем волокна предыдущей группы. Мышцы, содержащие эти волокна, развивают быстрое и сильное сокращение, но сравнительно быстро утомляются. Миоглобин в данной группе мышечных волокон отсутствует, вследствие чего мышцы, состоящие из волокон этого типа, называют белыми. Для мышечных волокон всех перечисленных групп характерно наличие одной, в крайнем случае, нескольких концевых пластинок, образованных одним двигательным аксоном. Тонические волокна. В отличие от предыдущих мышечных волокон в тонических волокнах двигательный аксон образует множество синаптических контактов с мембраной мышечного волокна. Развитие сокращения происходит медленно, что обусловлено низкой активностью миозиновой АТФ-азы. Также медленно происходит и расслабление. Мышечные волокна данного типа эффективно работают в изометрическом режиме. Эти мышечные волокна не генерируют потенциал действия и не подчиняются закону «все или ничего». Одиночный пресинаптический импульс вызывает незначительное сокращение. Серия импульсов вызовет суммацию постсинаптического потенциала и плавно возрастающую деполяризацию мышечного волокна. У человека мышечные волокна этого типа входят в состав наружных мышц глаза. Между структурой и функцией мышечных волокон существует тесная связь. Показано, что быстрые фазические волокна имеют высоко развитую саркоплазматическую сеть и обширную систему Т-трубочек, а медленные волокна – менее развитые СПР и сеть Т-системы. Кроме того, существует различие в активности кальциевых насосов саркоплазматической сети: в быстрых волокнах она значительно выше, что позволяет этим мышечным волокнам быстро расслабляться. Мышечные волокна не являются функциональной единицей скелетной мускулатуры. Эту роль выполняет нейромоторная, или двигательная единица, которая включает мотонейрон и группу мышечных волокон, иннервируемых разветвлениями аксона этого мотонейрона, расположенного в ЦНС. Число мышечных волокон, входящих в состав двигательной единицы, различно и зависит от функции, которую выполняет мышца в целом (табл.1). Таблица 1. Число мышечных волокон в двигательной единице различных мышц
В мышцах, обеспечивающих наиболее точные и быстрые движения, двигательная единица состоит из нескольких мышечных волокон, в то время как в мышцах, участвующих в поддержании позы, двигательные единицы включают несколько сотен и даже тысяч мышечных волокон. Величина потенциала покоя мышечных волокон составляет примерно — 90 мВ, потенциала действия — 120—130 мВ. Длительность потенциала действия 1—3 мс, величина критического потенциала — 50 мВ. Функции и свойства скелетных мышц Скелетная мускулатура является составной частью опорно-двигательного аппарата человека. При этом мышцы выполняют следующие функции: 1) обеспечивают определенную позу тела человека; 2) перемещают тело в пространстве; 3) перемещают отдельные части тела относительно друг друга; 4 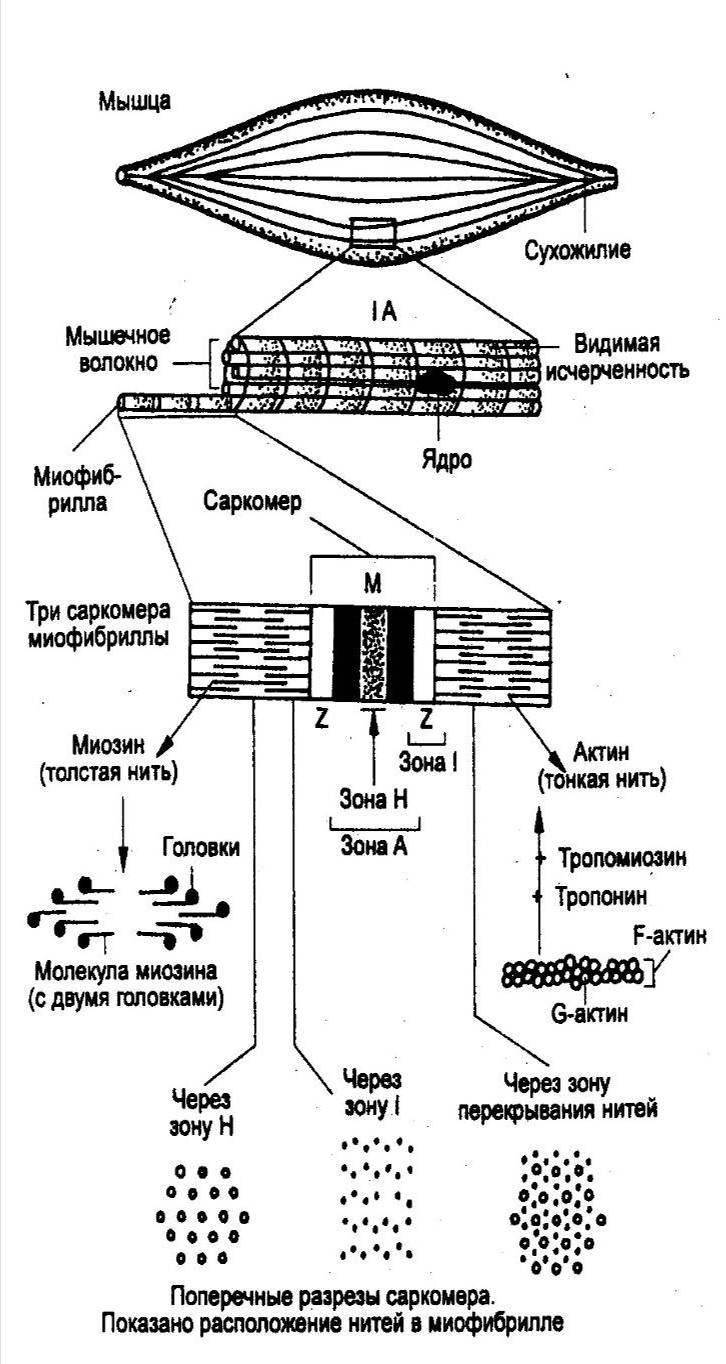 ) являются источником тепла, выполняя терморегуляционную функцию. ) являются источником тепла, выполняя терморегуляционную функцию.Скелетная мышца обладает следующими важнейшими свойствами: 1) возбудимостью — способностью отвечать на действие раздражителя изменением ионной проводимости и мембранного потенциала. В естественных условиях этим раздражителем является медиатор ацетилхолин, который выделяется в пресинаптических окончаниях аксонов мотонейронов. В лабораторных условиях часто используют электрическую стимуляцию мышцы. При электрической стимуляции мышцы первоначально возбуждаются нервные волокна, которые выделяют ацетилхолин, т. е. в данном случае наблюдается непрямое раздражение мышцы. Это обусловлено тем, что возбудимость нервных волокон выше мышечных. Для прямого раздражения мышцы необходимо применять миорелаксанты — вещества, блокирующие передачу нервного импульса через нервно-мышечный синапс; 2) проводимостью — способностью проводить потенциал действия по мембране вдоль и в глубь мышечного волокна по Т-трубочкам; 3) сократимостью — способностью укорачиваться или развивать напряжение при возбуждении; 4) эластичностью — способностью развивать напряжение при растягивании. Структурная организация мышечного волокна Мышечное волокно - это многоядерная структура, окружена мембраной и содержит сократительный аппарат — миофибриллы. В саркоплазме клеток содержатся митохондрии, системы продольных трубочек — саркоплазматическая сеть (ретикулум) и система поперечных трубочек — Т-система. Функциональной сократительной единицей мышечной клетки является саркомер (рис. 17); из саркомеров состоит миофибрилла. Саркомеры отделяются друг от друга Z-пластинками. Они последовательно расположены в миофибрилле, поэтому сокращение саркомеров вызывает сокращение миофибриллы и общее укорочение мышечного волокна. При изучении мышечных волокон в световой микроскоп выявило их поперечную исчерченность. Более тонкое, электронно-микроскопическое исследование показало, что поперечная исчерченность связана с особой организацией двух сократительных белков миофибрилл — актина (молекулярная масса 42 000) и миозина (молекулярная масса около 500000). Актиновые филаменты представлены двойной нитью, закрученной в двойную спираль с шагом около 36,5 нм. Их длина 1 мкм и диаметр 6—8 нм, одним концом прикреплены к Z-пластинке, количество актиновых нитей достигает около 2000. Между нитями актина в центре саркомера располагаются толстые нити миозина длиной около 1,6 мкм. В световом микроскопе это выглядит как полоска темного цвета (вследствие двойного лучепреломления) — анизотропный А-диск. В середине его видна более светлая полоска Н, в ней в состоянии покоя нет актиновых нитей. По обе стороны темного диска видны светлые изотропные полоски — I-диски, образованные нитями актина. В состоянии покоя нити актина и миозина незначительно перекрывают друг друга таким образом, что общая длина саркомера составляет около 2,5 мкм. Нити миозина в состоянии покоя удерживает М-линия, которая располагается в центре Н-полоски. На боковых сторонах миозиновых нитей были обнаружены поперечные мостики (рис.18). Они состоят из головки и шейки. Головка представляет собой шарнир и может поворачиваться вокруг своей оси за счет эластических свойств. Головка при связывании с актином приобретает выраженную АТФ-азную активность. На поперечном срезе мышечного волокна можно увидеть гексагональную организацию миофиламента: каждая нить миозина окружена шестью нитями актина (рис. 17). В продольных бороздках актиновых филаментов располагаются нитевидные молекулы белка тропомиозина. С шагом, равным 40 нм, к молекуле тропомиозина прикреплена молекула другого белка — тропонина (рис.18). Тропонин и тропомиозин являются регуляторными белками и играют важную роль в механизмах взаимодействия актина и миозина при сокращении. Механизм мышечного сокращения Скелетная мышца - это сложная система, трансформирующая хими-ческую энергию в механическую работу и тепло. В настоящее время хорошо исследованы молекулярные механизмы этого преобразования. В  процессе сокращения мышечного волокна в нем происходят следующие этапы: процессе сокращения мышечного волокна в нем происходят следующие этапы:1. Электрохимическое преобразование: а) генерация ПД. Рис. 18. Действие Ca2+ во время активации миофибриллы (В.М. Смирнов) А — актиновая и миозиновая нити на продольном сечении волокна; Б — они же на поперечном сечении. 1— «шейка», 2— «головка», 3 — актиновый мономер, 4 — тропонин, 5 — тропомиозин, 6 — миозиновая нить. б) распространение ПД по системе Т-трубочек, которая связывает поверхностную мембрану и сократительный аппарат мышечного волокна. в) распространение ПД по мембране Т-трубочек и саркоплазматического ретикулума приводит к активации ферментов, вследствие чего образуется инозитолтрифосфат (ИТФ). Он активирует кальциевые каналы и из мембран цистерн СПР высвобождаются ионы Са2+. Внутриклеточная концентрация их повышается. 2. Хемомеханическое преобразование: а) связывание ионов Са2+ с тропонином. Он меняет свою конформацию и смещается в желобок между нитями актина, при этом освобождаются активные центры на актиновых филаментах. После этого возможно взаимодействие поперечных мостиков миозина с актиновыми нитями. б) взаимодействие миозиновой головки с актином. При этом головка миозина присоединяется к первому из нескольких последовательно расположенных стабильных центров. Вращение головки приводит к увеличению упругой эластической тяги шейки поперечного мостика и увеличению напряжения. Головки поперечных мостиков последовательно соединяются и разъединяются с актиновыми филаментами, что приводит к плавному смещению тонких и толстых нитей. в) скольжение нитей актина и миозина относительно друг друга и уменьшение размеров саркомера и общей длины мышцы. Это приводит к развитию напряжения или укорочения мышечного волокна. В каждом цикле соединения и разъединения головки миозина с актином расщепляется одна молекула АТФ на каждый мостик. Необходимо подчеркнуть, что АТФ играет в мышечной работе двоякую роль: фосфорилируя миозин, он обеспечивает энергией сокращение, но, находясь в свободном состоянии, он обеспечивает и расслабление мышцы (ее пластификацию). При исчезновении АТФ из миоплазмы развивается непрерывное сокращение — контрактура. Для расслабления в первую очередь необходимо понижение концентрации ионов Са2+ в саркоплазме. Для этого существует специальный механизм — кальциевый насос, который активно возвращает кальций в цистерны. Он активируется неорганическим фосфатом, который образуется при гидролизе АТФ и обеспечивается энергией для работы освобождения ее при распаде АТФ (рис.19). Таким образом, АТФ является вторым важнейшим фактором, абсолютно необходимым для процесса расслабления. Некоторое время после смерти мышцы остаются мягкими вследствие прекращения тонического влияния мотонейронов. Затем концентрация АТФ снижается ниже критического уровня и возможность разъединения головки миозина с актиновым филаментом исчезает. Возникает явление трупного окоченения с выраженной ригидностью скелетных мышц. Режимы мышечных сокращений Режим сокращения мышечных волокон (МВ) определяется частотой раздражителей или импульсов, поступающих от мотонейронов. Так, если на икроножную мышцу лягушки наносить стимул или раздражения с интервалом, превышающим длительность одиночного сокращения (свыше 0,1с), то мышца в промежутке между двумя раздражителями успевает расслабиться и возникает одиночное сокращение (рис.20). Оно состоит из нескольких фаз: 1) латентный период – это время от момента нанесения раздражителя до начала укорочения МВ и обусловлен возбуждением мембраны мышечного волокна, распространением ПД по Т-трубочкам, образованием инозитолтрифосфата, повышением концентрации внутриклеточного кальция и активации поперечных мостиков. Он составляет около 2 мс (0,02с). 2 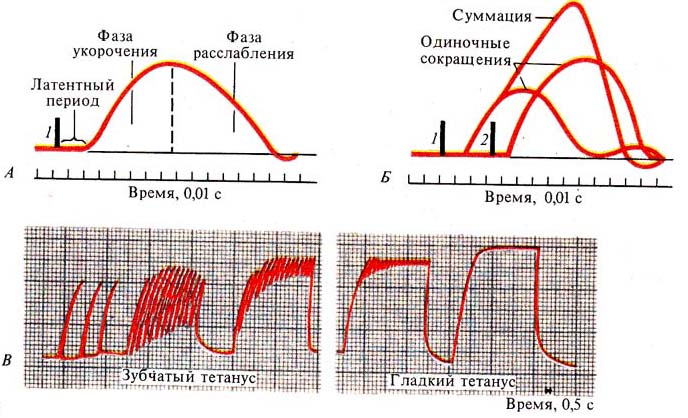 ) период укорочения или развития напряжения. Он длится 0,05 мс. В случае свободного укорочения мышечного волокна говорят об изотоническом режиме сокращения, при котором меняется только длина мышечного волокна, а напряжение остается относительно постоянным. Если мышечное волокно закреплено с двух сторон и не может свободно укорачиваться, то говорят об изометрическом режиме сокращения. В этом случае МВ укоротиться больше не может, а возникающее напряжение передается на эластические элементы (поперечные мостики миозиновых нитей, актиновые нити, Z-пластинки, продольно расположенная саркоплазматическая сеть и сарколемма мышечного волокна), расположенные внутри волокна. ) период укорочения или развития напряжения. Он длится 0,05 мс. В случае свободного укорочения мышечного волокна говорят об изотоническом режиме сокращения, при котором меняется только длина мышечного волокна, а напряжение остается относительно постоянным. Если мышечное волокно закреплено с двух сторон и не может свободно укорачиваться, то говорят об изометрическом режиме сокращения. В этом случае МВ укоротиться больше не может, а возникающее напряжение передается на эластические элементы (поперечные мостики миозиновых нитей, актиновые нити, Z-пластинки, продольно расположенная саркоплазматическая сеть и сарколемма мышечного волокна), расположенные внутри волокна. В процессе жизнедеятельности в организме человека развивается комбинация изотонического и изометрического компонентов - ауксотонический режим сокращения. Как правило, укорочение мышцы сопровождается развитием напряжения и увеличением силы, достаточной чтобы совершить перемещение груза. 3) период расслабления (0,07 мс), когда уменьшается концентрация ионов Са2+ и отсоединяются головки миозина от актиновых филаментов. П 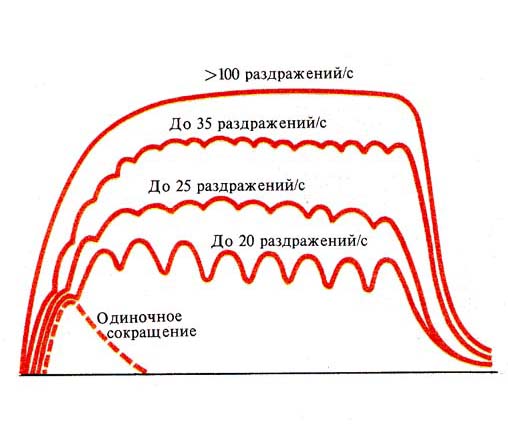 ри действии ритмических раздражителей развиваются тетанические сокращения. В зависимости от частоты раздражителя наблюдается возникновение зубчатого и гладкого тетануса (рис.21). При частоте стимулов до 20 Гц каждое следующее сокращение возникает в фазу расслабления от предыдущего и развивается зубчатый тетанус. При этом каждый последующий стимул будет воздействовать на мышцу до того, как она успеет полностью расслабиться после предыдущего раздражения. Возникает длительное слитное сокращение, в котором фаза расслабления оказывается незавершенной. На нее накладывается последующая фаза укорочения; амплитуды их с каждым стимулом суммируются и возрастают (зубчатый или неполный тетанус). ри действии ритмических раздражителей развиваются тетанические сокращения. В зависимости от частоты раздражителя наблюдается возникновение зубчатого и гладкого тетануса (рис.21). При частоте стимулов до 20 Гц каждое следующее сокращение возникает в фазу расслабления от предыдущего и развивается зубчатый тетанус. При этом каждый последующий стимул будет воздействовать на мышцу до того, как она успеет полностью расслабиться после предыдущего раздражения. Возникает длительное слитное сокращение, в котором фаза расслабления оказывается незавершенной. На нее накладывается последующая фаза укорочения; амплитуды их с каждым стимулом суммируются и возрастают (зубчатый или неполный тетанус).Увеличение частоты более 20 Гц отражает полную суммацию мышечных сокращений и наблюдается гладкий тетанус. В этом случае каждое следующее сокращение возникает в фазу укорочения или на высоте предыдущего сокращения. Так, как каждое последующее раздражение действует раньше, чем закончиться фаза укорочения предыдущего сокращения, то возникает гладкий, или полный тетанус, который характеризуется непрерывным сокращением, большим по сравнению с одиночными амплитудами сокращений и имеющим большее напряжение, развиваемое целой мышцей. Механизм суммаций сокращений при тетанусеАмплитуда тетанического сокращения мышцы превышает высоту ее одиночного сокращения. Г. Гельмгольц (1847) назвал этот процесс суперпозицией, то есть наложением сокращений, полагая, что эффект двух следующих друг за другом раздражений равен алгебраической сумме одиночных сокращений. Однако эти данные не соответствовали действительности. Н.Е. Введенский (1886) провел опыт, раздражая мышечное волокно пороговым раздражителем, возникало сокращение, дальнейшее раздражение подпороговыми раздражителями поддерживало амплитуду сокращения на первоначальном уровне. Н.Е Введенский объяснил это тем, что каждая последующая вспышка возбуждения и сокращения оставляет след в ткани в виде повышенной возбудимости. И действие раздражителя в момент, когда возбудимость мышцы повышена, амплитуда второго, а при ритмическом раздражении амплитуда тетанического сокращения становится больше, чем одиночного. В настоящее время установлена зависимость амплитуды тетанических сокращений от фазы возбудимости, в которую он попадает. Если второй стимул действует в период рефрактерности мышечного волокна, то он не вызовет повторного мышечного сокращения. Если же второй стимул действует на мышцу в период укорочения или начальную фазу расслабления, что совпадает с фазой экзальтации, то возникает тетаническое мышечное сокращение. Полагают, что в основе увеличения силы сокращений при действии ритмических раздражителей лежит повышение концентрации кальция внутри клетки, что позволяет осуществляться реакции взаимодействия актина и миозина и генерации мышечной силы поперечными мостиками достаточно длительное время. Утомление мышц. Причины утомления изолированной мышцы, нервно-мышечного препарата, утомления в естественных условиях Утомлением называется временное понижение работоспособности клетки, органа или целого организма, наступающее в результате работы и исчезающее после отдыха. Если длительно раздражать ритмическими электрическими стимулами изолированную мышцу, к которой подвешен груз, то амплитуда ее сокращений постепенно убывает, пока не дойдет до нуля. Полученная таким образом кривая называется кривой утомления. Наряду с изменением амплитуды сокращения при утомлении нарастает латентный период сокращения и увеличиваются пороги раздражения и хронаксия, то есть понижается возбудимость. Эти изменение возникают не сразу после работы, а спустя некоторое время, в течение которого наблюдается увеличение амплитуды одиночных сокращений мышцы. Этот период называется периодом врабатывания. При дальнейшем длительном раздражении развивается утомление мышечных волокон. Понижение работоспособности изолированной из организма мышцы при ее длительном раздражении обусловлено двумя основными причинами: первой из них является то, что во время сокращений в мышце накапливаются продукты обмена веществ (в частности, молочная, фосфорная кислоты и т. д.), оказывающие угнетающее влияние на работоспособность мышцы. Часть этих продуктов, а также ионы калия диффундируют из волокон наружу в околоклеточное пространство и оказывают угнетающее влияние на способность возбудимой мембраны генерировать потенциалы действия. Если изолированную мышцу, помещенную в раствор Рингера, довести длительным раздражением до полного утомления, то достаточно только сменить омывающую ее жидкость, чтобы восстановить сокращения мышцы. Другой причиной развития утомления изолированной мышцы является постепенное истощение в ней энергетический запасов. При длительной работе изолированной мышцы происходит резкое уменьшение запасов гликогена, вследствие чего нарушаются процессы ресинтеза АТФ и креатинфосфата, необходимых для осуществления сокращения. Утомление нервно-мышечного препарата обусловлено следующими причинами. При длительном раздражении нерва нарушение нервно-мышечной передачи развивается задолго до того, как мышца, а тем более нерв в силу утомления утрачивает способность к проведению возбуждения. Объясняется это тем, что в нервных окончаниях при длительном раздражении уменьшается запас "заготовленного" медиатора. Поэтому порции ацетилхолина, выделяющиеся в синапсах в ответ на каждый импульс, уменьшаются и постсинаптические потенциалы снижаются до подпороговых величин. Наряду с этим при длительном раздражении нерва происходит постепенное понижение чувствительности постсинаптической мембраны мышечного волокна к ацетилхолину. В результате уменьшается величина потенциалов концевой пластинки. Когда их амплитуда падает ниже некоторого критического уровня, возникновение потенциалов действия в мышечном волокне прекращается. По этим причинам синапсы быстрее утомляются, чем нервные волокна и мышцы. Следует отметить, что нервные волокна обладают относительной неутомляемостью. Впервые Н.Е. Введенский показал, что нерв в атмосфере воздуха сохраняет способность к проведению возбуждений даже при многочасовом непрерывном раздражении (около 8 часов). Относительная неутомляемость нерва отчасти зависит от того, что нерв тратит при своем возбуждении сравнительно мало энергии. Благодаря этому процессы ресинтеза в нерве в состоянии покрывать его относительно малые расходы при возбуждении даже в том случае, если это возбуждение длится много часов. Необходимо отметить, что утомление изолированной скелетной мышцы при ее прямом раздражении является лабораторным феноменом. В естественных условиях утомление двигательного аппарата при длительной работе развивается более сложно и зависит от большего числа факторов. 1. В организме мышца непрерывно снабжается кровью, и, следовательно, получает с ней определенное количество питательных веществ (глюкоза, аминокислоты) и освобождается от продуктов обмена, нарушающих нормальную жизнедеятельность мышечных волокон. 2. В целом организме утомление зависит не только от процессов в мышце, но и от процессов, развивающихся в нервной системе, участвующих в управление двигательной деятельностью. Так, например, утомление сопровождается дискоординацией движений, возбуждением многих мышц, которые не участвуют в совершении работы. Активный отдых и его механизм. (И.М. Сеченов, феномен Орбели-Гинецинского) При выявлении причин утомления двигательного аппарата в применении к целостному организму в настоящее время нередко различают два вида двигательной деятельности: локальную, когда активно сравнительно небольшое количество мышц, и общую, когда в работе участвует большинство мышц тела. В первом случае среди причин утомления на первое место выступают периферические факторы, то есть процессы в самой мышце; во втором ведущее значение приобретают центральные факторы (нервная система) и недостаточность вегетативного обеспечения движений (дыхания, кровообращения). Впервые И.М. Сеченов (1903) показал, что восстановление работоспособности утомленных мышц руки человека после длительной работы по подъему груза резко ускоряется, если в период отдыха производить работу другой рукой. Временное восстановление работоспособности мышц утомленной руки может быть достигнуто и при других видах двигательной активности, например, при работе различных мышц нижних конечностей. В отличие от простого покоя такой отдых был назван И.М. Сеченовым активным. Сеченов рассматривал эти факты как доказательство того, что утомление, прежде всего, развивается в нервных центрах. Убедительным доказательством роли изменения состояния нервных центров в развитии утомления в целом организме служат опыты с внушением. Так, исследуемый может длительно поднимать тяжелую гирю, если ему внушить, что в его руке находится легкая корзина. Напротив, если внушить исследуемому, поднимающему легкую корзину, что ему дана тяжелая гиря, то быстро развивается утомление. При этом изменение пульса, дыхания и газообмена находится в соответствии не с реальной, осуществляемой человеком работой, а с той, которая ему внушена (В.М. Василевский, Д.И. Шатенштейн). Феномен Орбели-Гинецинского был открыт в 1923 г. В опытах на нервно-мышечном препарате двигательные волокна раздражались электростимулятором. Изолированная мышца отвечала сокращением на каждое из ритмически повторяющихся раздражений, и на ленте кимографа регистрировалась типичная кривая мышечного сокращения. По мере утомления амплитуда кривой снижалась. После раздражения симпатических нервов, происходило увеличение амплитуды сокращений мышцы, и на кимограмме отмечалась новая волна повышенной активности. Позднее феномен был подтвержден и на мышцах млекопитающих в условиях нормального кровоснабжения. Л.А. Орбели выдвинул представление об универсальной адаптационно-трофической функции симпатической нервной системы, регулирующей функциональные свойства всех органов и тканей, устанавливая их на оптимальный для данных условий уровень. Эта регуляция не ограничивается гладкими мышцами и железами, она охватывает все звенья рефлекторной дуги - рецепторы, центральную нервную систему, нервные проводники и скелетную мускулатуру. В основе феномена Орбели-Гинецинского лежит активация симпатической нервной системы. Дальнейшие исследования позволили выявить общность влияния симпатической нервной системы и ретикулярной формации головного мозга на восстановление работоспособности мышц. Рабочая гипертрофия и атрофия от бездеятельности Систематическая интенсивная работа мышц приводит к увеличению массы мышечной ткани. Это явление носит название рабочей гипертрофии мышцы. Рабочая гипертрофия мышцы происходит отчасти за счет продольного расщепления, а главным образом за счет утолщения (увеличения диаметра) мышечных волокон. Можно выделить два основных типа рабочей гипертрофии мышечных волокон. Первый тип - саркоплазматический - утолщение мышечных волокон за счет преимущественного увеличения объема саркоплазмы, то есть несократительной части мышечных волокон. Этот тип гипертрофии приводит к повышению метаболических резервов мышцы: гликогена, безазотистых веществ, креатинфосфата, миоглобина и др. Значительное увеличение числа капилляров в результате тренировки также может в какой-то мере вызывать утолщение мышцы. Первый тип рабочей гипертрофии мало влияет на рост силы мышц, но зато значительно повышает способность их к продолжительной работе, то есть выносливость. Второй тип рабочей гипертрофии - миофибриллярный - связан с увеличением объема миофибрилл, то есть собственно сократительного аппарата мышечных волокон. При этом мышечный поперечник может увеличиваться не очень значительно, так как в основном возрастает плотность укладки миофибрилл в мышечном волокне. Второй тип рабочей гипертрофии ведет к значительному росту максимальной силы мышцы. Существенно увеличивается и абсолютная сила мышцы, тогда как при первом типе рабочей гипертрофии она или совсем не изменяется, или даже несколько уменьшается. Преимущественное развитие первого или второго типа рабочей гипертрофии определяется характером мышечной тренировки. Вероятно, длительные динамические упражнения с относительно небольшой нагрузкой вызывают рабочую гипертрофию главным образом первого типа (преимущественное увеличение объема саркоплазмы, а не миофибрилл). Изометрические упражнения с применением больших мышечных напряжений (более 2/3 от максимальной произвольной силы тренируемых мышечных групп), наоборот способствуют развитию рабочей гипертрофии второго типа (миофибриллярной гипертрофии). В основе рабочей гипертрофии лежит интенсивный синтез мышечных белков, ДНК и РНК. Очень важную роль в регуляции объема мышечной массы играют гормоны - андрогены. У тренированных людей, у которых многие мышцы гипертрофированы, мускулатура может составлять до 50% массы тела (вместо 35-40% в норме). Противоположным рабочей гипертрофии состоянием является атрофия мышц от бездеятельности. Она развивается во всех случаях, когда мышца почему-либо длительно не совершает нормальной работы. Это наблюдается, например, при обездвижении конечности в гипсовой повязке, долгом пребывании больного в постели, перерезке сухожилия, вследствие чего мышца перестает совершать работу. При атрофии диаметр мышечных волокон и содержание в них сократительных белков, гликогена, АТФ и других, важных для сократительной деятельности веществ, уменьшаются. После возобновления нормальной работы атрофия мышцы постепенно исчезает. Действие электрического тока на возбудимые ткани Влияние длительного постоянного тока При замыкании электрической цепи в I фазу длительного действия постоянного тока под катодом наблюдается снижение мембранного потенциала на 3-5 мВ (деполяризация) и возбудимость увеличивается. Под анодом - мембранный потенциал возрастает на 3-5 мВ (гиперполяризация) и возбудимость снижается. Этот пассивный сдвиг потенциала говорит об электротонических изменениях или электротоне (рис.23). Под катодом развивается катэлектротон, при котором наблюдается снижение мембранного потенциала, увеличение белкового обмена, повышение проницаемости для ионов натрия, а под анодом, наоборот, повышается мембранный потенциал, уменьшается белковый обмен, снижается проницаемость для ионов натрия - анэлектротон. Во II фазу к сдвигам мембранного потенциала присоединяется и изменение критического потенциала и развиваются катодическая депрессия и анодическая экзальтация (рис.23). Катодическая депрессия. Под катодом развивается смещение уровня критической деполяризации вверх (рис.24), за счет инактивации натриевых каналов, и возбудимость снижается. Это явление называется аккомодацией. В результате этого в клетке развиваются низкоамплитудные потенциалы действия и ответная реакция на действие раздражителя снижается. Анодическая экзальтация. Под анодом вследствие длительного действия электротока уровень критического потенциала смещается вниз (рис.18). При этом уменьшается порог деполяризации и возбудимость увеличивается вследствие повышения натриевой проницаемости. Влияние кратковременного постоянного тока Еще в 1859г. Пфлюгер показал, что при кратковременном действии электротока на клетку возбуждение возникает только в момент замыкания цепи под катодом и в момент размыкания цепи под анодом (рис.18). Это получило название – закона полярного действия. Объясняется он тем, что под катодом при замыкании цепи возникает частичная деполяризация мембраны клетки, так как катод электроотрицателен. А в области действия анода заряд клетки наоборот гиперполяризуется и возбуждения при замыкании 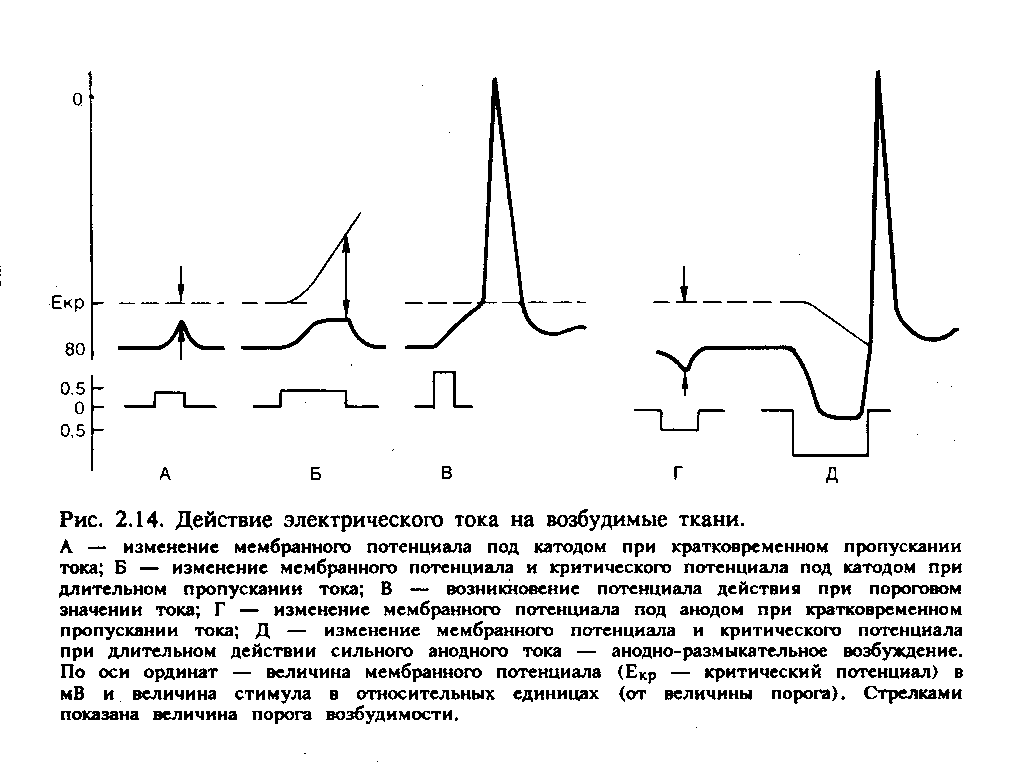 не возникает. Это связано с тем, что при действии анода уровень критической деполяризации (Екр) сдвигается в сторону поляризации клетки, а при размыкании цепи мембранный потенциал возвращается к исходной величине и достигает Екр, вследствие чего возникает возбуждение (рис.24). Особенности физиологии возбудимых тканей у детей Особенности физиологии нервов Проводимость новорожденного ребенка ниже, чем у взрослого в два раза и скорость проведения возбуждения составляет около 50% от таковой у взрослых. Проведения возбуждения по нервным волокнам плохо изолировано. В процессе взросления нервные волокна миелинизируются, увеличивается диаметр осевого цилиндра и волокна в целом, а чем толще становится волокно, тем меньше продольное сопротивление ионному току. Это приводит к тому, что скорость распространения ПД увеличивается. У детей он достигает показателей взрослого человека к 5-9 годам для разных нервных волокон. Так, передние спинномозговые корешки созревают к 2-5 году жизни, а задние спинномозговые корешки – к 5-9 году. Возбудимость нервных волокон новорожденного значительно ниже, чем у взрослого. Характеристикой этого является хронаксия, величина которой в несколько раз выше; потенциал покоя, который у детей значительно ниже. Низкая величина потенциала покоя связана с тем, что клеточная мембрана имеет высокую ионную проницаемость и постоянно происходит утечка ионных токов. Это приводит к снижению трансмембранной разности ионов (градиенту концентрации) и приводит к формированию низкой амплитуды потенциала действия в сочетании с большей его продолжительностью и отсутствием реверсии. В процессе роста проницаемость мембраны уменьшается и мембранный потенциал достигает показателей взрослого человека. Соответственно увеличивается и амплитуда потенциала действия, скорость проведения ПД возрастает, так как при высокой амплитуде легче вызвать возбуждение соседнего участка волокна. У плода и ребенка первых лет жизни мякотные волокна плохо миелинизированы и каналы для натрия и калия располагаются равномерно. В онтогенезе волокно миелинизируется, ионные каналы концентрируются в перехватах Ранвье, расстояние между перехватами увеличивается. Это характеризует структурную зрелость мякотных волокон. В безмякотных волокнах распределение ионных каналов остается равномерным. Лабильность нервных волокон новорожденных также низка. У детей более старшего возраста она увеличивается за счет снижения длительности рефрактерного периода и увеличения скорости проведения возбуждения. Особенности физиологии мышц Возбудимость мышц новорожденного очень низка. Показателем этого является высокая хронаксия и большой порог деполяризации. У новорожденного МП миоцитов составляет -20-40мВ. Трансмембранная разность ионов К+ и Nа+ невысока. Поэтому и величина ПД также небольшая. Кроме того, отмечается длительность фаз абсолютной и относительной рефрактерности. В процессе роста проницаемость мембраны уменьшается, работа ионных насосов улучшается и увеличивается МП и ПД. Лабильность детей ниже, чем у взрослых вследствие большой длительности рефрактерных фаз. с возрастом происходит укорочение фаз абсолютной и относительной рефрактерности и как следствие возрастание скорости проведения возбуждения и увеличению быстроты движений. Проводимость. Низкая у новорожденных скорость проведения ПД, с возрастом увеличивается. К этому приводит увеличение толщины мышечного волокна и увеличение амплитуды потенциала действия, так как снижается сопротивление ионному току и быстрее развивается возбуждение на соседнем участке мембраны. Сократимость. Одиночные сокращения мышц новорожденного замедленны – как фаза укорочения, так и фаза расслабления - и характеризуется большим временем сокращения. В мышцах ребенка быстрее накапливаются продукты метаболизма и, поэтому, тетанус имеет пологое начало и постепенное расслабление, как тетанус утомленной мышцы. Мышцы отвечают тоническим сокращением на раздражители любой частоты и сокращаются без пессимального торможения столько, сколько действует раздражитель. Это связано с недостаточной зрелостью мионевральных синапсов. У новорожденных отсутствует разделение мышц на быстрые и медленные, но уже с первых дней жизни у ребенка начинается постепенная дифференцировка, характерная для взрослых. Эластичность мышц новорожденного выше, чем у взрослого и с возрастом уменьшается. А упругость и прочность, наоборот, увеличиваются. |