Лекция 1-4. Лекция 1 Физиология, ее предмет, роль в системе медицинского образования и задачи
 Скачать 0.84 Mb. Скачать 0.84 Mb.
|

Методы физиологических исследований1. Органолептический метод (при помощи органов осязания) - включает осмотр кожных покровов, склер глаз, слизистых оболочек, перкуссию, аускультацию, пальпация. Перкуссия (выстукивание) - метод исследования внутренних органов, основанный на выстукивании по поверхности тела обследуемого с оценкой характера возникающих при этом звуков. Этот метод позволяет определить границы органов, их величину и форму. изменения звука свидетельствует о наличие патологии (при заболеваниях легких) Аускультация (выслушивание) - метод исследования внутренних органов, основанный на выслушивании звуковых явлений, связанных с их деятельностью. Производится при помощи стетоскопа и фонендоскопа. Определяются типы дыхания, тоны сердца и частота сердечной деятельности, сердцебиение плода. Пальпация (ощупывание) - клинический метод исследования при помощи осязания с целью изучения физических свойств и чувствительности тканей и органов, топографических соотношений между ними и обнаружения некоторых функциональных явлений в организме (температуры кожи, пульсации сосудов, перистальтики кишечника и др.). 2. Инструментальный метод. С развитием техники увеличивалось количество диагностического оборудования. Инструментальный метод основан на изучении функций организма при помощи различной медицинской аппаратуры (электрокардиография, фонокардиография, рентгенография, ультразвуковое исследование, миография, эндоскопия и т.д.). 3. Морфологический метод - изучение состояния клеток. Например, биопсия, цитологическое исследование. 4. Клинико-биохимический метод - исследование биологических жидкостей организма. Сюда относится клиническое и биохимическое исследование слюны, крови, лимфы, спинномозговой жидкости и т.д. 5. Паталогоанатомический метод - изучение изменений во внутренних органах после смерти. 6. Сбор анамнеза - анамнеза жизни и анамнеза болезни. Анамнез болезни отражает развитие болезни от ее начала до настоящего момента. Анамнез жизни представляет собой медицинскую биографию больного по основным периодам его жизни (младенчество, детство, юношество, зрелый возраст). Кроме перечисленных методов исследования заслуживает особого внимания экспериментальный метод, который включает метод острого и хронического эксперимента. Метод острого эксперимента. В основе метода лежит хирургическое вмешательство (вивисекция, живосечение) для того чтобы получить доступ к какому-либо внутреннему органу или системе (изучение сердечной деятельности, возбудимости мышечных и нервных волокон и т.д.). У этого метода много недостатков, искажающих результаты: 1. увеличение стрессорных и болевых веществ, которые выделяются при проведении операции, 2. наличие наркотического вещества, 3. операция, 4. кровопотеря. Все это совершенно меняет и нарушает нормальное течение физиологических процессов, для изучения которых и предпринимался этот опыт. Кроме того, исследование изолированных органов не давало представления об их истинной функции в условиях целостного неповрежденного организма. Метод хронического эксперимента. И.П. Павлов нашел способ, позволяющий рассмотреть функции организма, не нарушая его целостности. Это был метод хронического эксперимента. На наркотизированном животном в условиях стерильности предварительно проводили сложную операцию, позволяющую получить доступ к внутренним органам: вживляли фистульную трубку или выводили наружу и подшивали к коже проток железы. Сам опыт начинался много дней спустя, когда рана заживала, животное выздоравливало и по характеру течения физиологических процессов практически ничем не отличалось от нормального, здорового. Благодаря наложенной фистуле можно было длительно изучать течение тех или иных физиологических процессов в естественных условиях. Биологические реакции Все биологические структуры отличаются способностью к раздражимости, то есть могут отвечать на внешние и внутренние воздействия изменением обмена веществ и морфо-функциональными реакциями. Раздражимость - это потенциальная способность биологических структур к изменению метаболизма под действием раздражителя. Само же изменение обмена веществ называется раздражением. За счет такого свойства как раздражимость все живые системы адаптируются к изменению окружающей среды, что способствует живым организмам выживать в изменчивом мире. С развитием у многоклеточных организмов нервной и мышечной систем, возникла более совершенная форма реагирования на раздражители - возбудимость, обеспечивающая наибольшую быстроту и точность ответных реакций. Возбуждение - это быстрый процесс, возникающий в результате действия раздражителя на живой объект как проявление свойства возбудимости. Такое свойство приобрели 3 ткани: нервная, мышечная и железистая. За счет появления возбудимых тканей организмы получили возможность реагировать на внешние воздействия не медленными, а срочными реакциями, что существенно улучшило их адаптацию к среде. Возбуждение характеризуется следующими признаками: 1. переход от состояния физиологического покоя в состояние возбуждения; 2. возникнув в месте действия раздражителя, возбуждение распространяется по всей клетке и ткани; 3. возбуждение намного кратковременнее, чем раздражение и затухает после прекращения действия раздражителя; 4. каждая возбудимая ткань генерирует специфический ответ: нервная ткань - генерация и проведение нервного импульса, мышечная ткань - сокращение, эпителиальная ткань - образование и выделение секрета. Ответная реакция на раздражение невозбудимых систем (костной, соединительнотканной, эпителиальной ткани, крови) заключается в физико-химической, обменной, а иногда и структурной перестройке. Обмен веществ никогда не возникает сам по себе, реакция раздражения не возникает без причины, для этого необходимы факторы, вызывающие возникновение ответной реакции. Они называются раздражителями. Раздражитель – это любая форма движения материи, любые агенты внешней и внутренней среды. Классификация раздражителей По виду энергии раздражители делятся на следующие группы: 1. Физические – к ним относятся температура, звук, механические факторы. Так снижение температуры (гипотермия до 18-20оС) ведет к ослаблению обмена веществ в тканях, что с успехом используется при операциях на сердце, при такой температуре мозг обходится без питания 10-15 мин. Чаще всего физиологи используют в качестве раздражителя электрический ток. Это связано с тем, что он по своему характеру ближе всего стоит к нервному импульсу – естественному раздражителю всех тканей и органов, так же он легко дозируется по силе, длительности и частоте стимулов. К механическим раздражителям относят удар, укол, прикосновение, вибрацию, растяжение, сдавление. Но они применяются реже. 2. Химические раздражители – кислоты, щелочи, соли существенно меняют режим обмена. Редко кто сможет задержать дыхание больше 1-2 мин., это связано с закислением крови, что приводит к усилению деятельности дыхательного центра. 3. Физико-химические раздражители – изменение осмотического давления, рН, коллоидного состояния. Осмотическое давление крови соответствует 0,9% раствору NaCl. Внутривенное введение гипотонических растворов вызывает серьезные нарушения: разрушение эритроцитов, внутрисосудистое свертывание крови. 4. Биологические раздражители – ими являются вещества, образующиеся в ходе обычного обмена веществ: мочевина, аминокислоты, продукты распада жиров, белков, углеводов. Они могут действовать местно, расширяя сосуды в работающей мышце, а также рефлекторным путем, т.е через нервную систему могут регулировать деятельность других органов и систем. К биологическим раздражителям относятся гормоны – вещества, образующиеся в эндокринных железах и обладающие чрезвычайно высокой активностью. Так 1г адреналина стимулирует работу 100 млн. изолированных сердец лягушек. 5. Все перечисленные выше раздражители являются общими для всех живых организмов. Кроме того, у человека существует специфический раздражитель, связанный с его жизнью в обществе – это социальный раздражитель. Одним из таких раздражителей является слово. Слово – очень сильный раздражитель, способный резко изменить нашу жизнь. Примером может являться внушение и гипноз. Зная о силе слова, врач должен действовать очень осторожно: словом можно не только помочь, но и вызвать тяжелые ятрогенные заболевания. По характеру действия раздражители делят на контактные и дистантные. Контактные действуют непосредственно на ткань, а у дистантных – источник раздражения располагается на расстоянии. По физиологическому значению все раздражители делятся на адекватные, специфические и неадекватные, неспецифические. Адекватными называются раздражители, к которым ткань, орган приспособился в процессе эволюции, Так для глаза адекватным раздражителем служит свет. При этом достаточно раздражителя небольшой силы, чтобы возникла ответная реакция, сетчатка глаза воспринимает свет силой всего в 2-4 кванта. Но также чувство света (звездочки в глазах) можно вызвать при сильном ударе по глазу. Это уже будет неадекватный раздражитель, сила которого значительно превышает силу адекватного раздражителя. Физиологи давно пытались количественно оценить возбудимость тканей. Для этой цели введено понятие о пороге силы раздражения. Пороговая сила - это минимальная величина раздражителя, способная вызвать ответную реакцию. Для лучшего понимания обратимся к примеру определения порога раздражения мышцы. Мышцу начинаем раздражать током силой в 3 мВ – это не вызывает сокращения, повышаем силу тока до 5 мВ, возникает слабое сокращение. В зависимости от этого все раздражители делят на пороговые, подпороговые и надпороговые. В нашем случае подпороговым раздражителем будет являться раздражитель силой в 3 мВ, а пороговым - раздражитель в 5мВ. Раздражитель должен действовать в течение некоторого времени, для того чтобы сформировалась ответная реакция. И было введено понятие порога времени раздражения - это то минимальное время, течение которого должен действовать раздражитель пороговой силы, для возникновения ответной реакции. Между силой и временем действия имеется прямая зависимость, чем больше порог силы, тем меньше порог времени. Так было установлено, что самой возбудимой является нервная ткань, на втором месте стоит мышечная и на последнем – железистая. Между порогом возбудимости и возбудимостью имеется обратная зависимость: чем меньше порог, тем выше возбудимость и наоборот. Сравним возбудимость двух мышц: одна сокращается при действии током в 5 мВ, а другая – 25 мВ. Ясно, что более возбудима первая, которая реагирует на более слабый раздражитель. Самый низкий порог раздражения имеет нервная ткань, затем - мышечная, и на последнем месте – железистая. Закон "все или ничего" Зависимость величины ответной реакции от силы раздражителя характеризуется несколькими законами раздражения: закон "все или ничего" и "правило силовых отношений". Закон "все или ничего" Согласно этому закону, подпороговые раздражения не вызывают возбуждения ("ничего"), при пороговых и надпороговых стимулах возбуждение сразу приобретает максимальную величину ("все") и уже не увеличивается при дальнейшем усилении раздражения. По этому закону функционируют структурные единицы – мышечное волокно, нервное волокно. Эта закономерность первоначально была открыта Г. Боудичем в 1876 году при исследовании сердца, а в дальнейшем подтверждена и на других возбудимых тканях. Так, в 1945 году Като провел эксперимент на изолированной системе. Он изолировал мышечное волокно и действовал раздражителями различной силы. В качестве раздражителя использовал электрический ток. При действии раздражителей подпороговой величины 1, 2, 3, 4 мВ мышечное волокно не сокращалось. Пороговый раздражитель силой 5 мВ вызывал сокращение изолированного волокна и дальнейшее увеличение силы тока не вызывало усиления амплитуды сокращения. 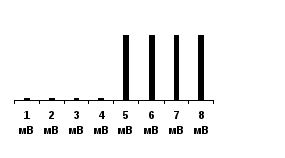 В живой клетке существуют процессы, активно приостанавливающие этот процесс. Если на нервное или мышечное волокно действовать веществами, уменьшающими проницаемость для ионов натрия (кокаин, новокаин, уретан), это приведет к понижению возбуждения в исследуемом волокне, и, наоборот, влияние стимулов, приводящих к уменьшению проницаемости для ионов калия, способствует увеличению амплитуды возбуждения. В живой клетке существуют процессы, активно приостанавливающие этот процесс. Если на нервное или мышечное волокно действовать веществами, уменьшающими проницаемость для ионов натрия (кокаин, новокаин, уретан), это приведет к понижению возбуждения в исследуемом волокне, и, наоборот, влияние стимулов, приводящих к уменьшению проницаемости для ионов калия, способствует увеличению амплитуды возбуждения.Закон силовых отношенийФункциональная единица (мышца, нерв) реагируют на изменение силы раздражителя по закону силовых отношений. Чем сильнее ток, тем сильнее реакция. Эта закономерность на раздражение выражается законом градуальности (силовых отношений). В 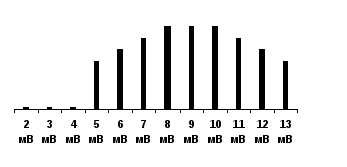 1876 году Боудич, а затем в 1905 году Люкас провел эксперимент на межпальцевой мышце лягушки, состоящей из 5 мышечных волокон. При действии подпороговых раздражителей (1, 2, 3 мВ) сокращения мышцы не происходило. Раздражитель пороговой силы (4 мВ) вызывал сокращение целой мышцы. Увеличение силы раздражителя (5, 6, 7, 8 мВ) вызывало увеличение амплитуды сокращения мышцы, пока сокращение не достигало максимальной величины (оптимум). После этого, дальнейшая стимуляция возрастающим по силе электрическим током (9, 10, 11, 12, 13 мВ) приводило к уменьшению амплитуды сокращения (пессимум). 1876 году Боудич, а затем в 1905 году Люкас провел эксперимент на межпальцевой мышце лягушки, состоящей из 5 мышечных волокон. При действии подпороговых раздражителей (1, 2, 3 мВ) сокращения мышцы не происходило. Раздражитель пороговой силы (4 мВ) вызывал сокращение целой мышцы. Увеличение силы раздражителя (5, 6, 7, 8 мВ) вызывало увеличение амплитуды сокращения мышцы, пока сокращение не достигало максимальной величины (оптимум). После этого, дальнейшая стимуляция возрастающим по силе электрическим током (9, 10, 11, 12, 13 мВ) приводило к уменьшению амплитуды сокращения (пессимум).Почему же это происходит? Природа закона силовых отношений. Мышца состоит из отдельных мышечных волокон, причем каждое волокно имеет различную возбудимость. При действии раздражителя пороговой силы возбуждаются более возбудимые мышечные волокна (пороговое сокращение). По мере увеличения силы тока в процесс возбуждения вовлекаются менее возбудимые мышечные волокна (резерв или субмаксимальные сокращения). В момент сокращения всех волокон регистрируется максимальное сокращение (предел). После этого наступает пессимум - ослабление реакции на очень сильное и очень частое возбуждение. Развивается тормозной ответ. Закон силовых отношений имеет несколько свойств: 1) каждая возбудимая ткань имеет свой функциональный резерв; 2) каждая возбудимая ткань имеет свой функциональный предел (оптимум). Лекция 2 Биотоки. Мембранный потенциал Как уже говорилось, процесс возбуждения в каждой возбудимой ткани проявляется специфической реакцией. В то же время процессу возбуждения во всех тканях присущи общие, универсальные признаки, связанные с особенностями метаболизма при возбуждении. При этом происходит значительное увеличение обмена веществ. Энергия химических реакций преобразуется в механическую энергию сокращения, секрецию, в тепло и небольшая часть превращается в электрические явления. В них переходит лишь 1/1 000 000 часть всей энергии возбуждения, но эти токи сейчас хорошо регистрируются. Электрические явления у животных известны давно. На первом месте по мощности электрического органа располагается электр.угорь из рек Южной Америки, длина этой рыбы 2м, а длина эл. органа 1,5м, мощность его разряда600в и частота 1600 в сек. Его удары мгновенно убивает крупных животных и человека, находящихся в воде. Следующим по мощности является электрический орган электр.ската из Атлантического океана, далее располагается электр. орган нильского сома, который иннервируется одной нервной клеткой. Именно на нем Бабухин доказал 2-сторонее проведение возбуждения по нервам. Электр.явления присущи не только рыбам. Они существуют во всех клетках каждого животного, где впервые были обнаружены в 1786 году Луиджи Гальвани, итальянским врачом и физиологом Болонского университета. Лягушачьи лапки он подвешивал за седалищный нерв на медный крючок, нижним концом они касались железных перил и происходило стойкое сокращение, как при раздражении электрическим током. Гальвани истолковал это как критерий наличия животного электричества, т.е. мышца и нерв имеют различные заряды, металл замыкает цепь между ними, приводя к сокращению (рисунок). Этот опыт получил название первого опыта Гальвани. Против утверждения Гальвани о животном электричестве выступил итальянский физик и физиолог Александро Вольта Он в 1791 году подтвердил опыт Гальвани, но дал иное объяснение: сокращение мышцы происходит из-за разности зарядов между разнородными металлами, а не между тканями. В 1793-94 годах Гальвани провел опыт «сокращения без металлов». При этом нерв набрасывался на мышцу с помощью непроводника - деревянной или стеклянной палочки, но мышца все равно сокращалась. Этот эксперимент в классике физиологии известен, как второй опыт Гальвани. Впоследствии оказалось, что оба ученых были правы: существует и животное электричество, и разность потенциалов между металлами. В честь Вольта животное электричество измеряется в вольтах, а ток между разными металлами, находящимися в электролите в честь Гальвани назвали гальваническим. В первой половине 19 века (в 1848г.) нем. физиолог Э.Дюбуа-Реймон впервые использовал в физиологии физические приборы (гальванометр) и измерил величину животного электричества. Он накладывал электроды на поврежденный и неповрежденный участок мышцы лягушки, при этом стрелка гальванометра отклонялась в сторону поврежденного участка. Из курса физики вы знаете, что в электрической цепи ток движется от «+» к «-», следовательно поврежденный участок заряжен отрицательно по отношению к неповрежденному, т.е внутри клетка заряжена отрицательно, а снаружи – положительно. Таким образом, в состоянии покоя существует трансмембранная разность потенциалов между цитоплазмой и окружающим клетку наружным раствором. Такая разность потенциалов, свойственная состоянию покоя называется потенциалом покоя или мембранным потенциалом. Кроме того, выделяют еще ток повреждения – разность потенциалов между поврежденным и неповрежденным участками, она заметно меньше мембранного потенциала, так как межклеточная жидкость шунтирует регистрирующую систему, поэтому точную величину МП удалось измерить недавно. Это удалось лишь в 1950 году, когда Линг и Джерард предложили стеклянные микроэлектроды, которые можно вводить в клетку не повреждая ее. Они представляют собой тонкие капилляры диаметром один микрон, заполненные 3М р-ром КСl. Один электрод вводят внутрь клетки, а второй накладывают на поверхность клетки. Пока микроэлектрод не проколол мембрану, осциллограф пишет прямую линию на уровне 0 – разности потенциалов нет, как только оболочка проколота, стрелка гальванометра сразу отклоняется вниз, регистрируя отрицательный заряд цитоплазмы. Возбудимые ткани имеют различную величину потенциала покоя: у скелетной мускулатуры он равен -80-90 мВ, у нервной ткани - -70 мВ, у гладкой мускулатуры - 50 - 60 мВ и у железистого эпителия - 20 мВ. МП поддерживается на примерно постоянном уровне на протяжении всей жизни и исчезает при гибели клетки. Еще в 1896 году В.Ю. Чаговец высказал гипотезу об ионном механизме электрических потенциалов в живых тканях и сделал попытку объяснить с позиции теории электролитической диссоциации Аррениуса. Сейчас природа мембранного потенциала объясняется с позиции мембранно-ионной теории Юлиуса Бернштейна. Он выдвинул ее в 1902, а подтверждена она была лишь в 1961 английскими физиологами А. Ходжкиным, Хаксли и Экклсом (за что в 1963 году ими была получена Нобелевская премия). Согласно этой теории для возникновения поляризации клеточной мембраны необходимы условие и причина поляризации клеточных мембран. Условием является наличие полупроницаемой мембраны, причиной - трансмембранная разность ионов по обе стороны мембраны. Вначале разберем строение и свойства мембраны. Строение и свойства мембраны Толщина клеточных мембран колеблется от 6 до 12 нм. Мембрана состоит из фосфолипидов и белков. В настоящее время получила распространение жидкостно-мозаичная модель строения мембран (рис.6). Она представляет собой двойной слой фосфолипидов, гидрофобные хвосты которых направлены внутрь мембраны, а гидрофильные головки ориентированны к ее поверхности. Глобулярные белки в зависимости от расположения называются периферическими, полуинтегральными и интегральными. Периферическими называются белки, которые располагаются на поверхности мембраны, полуинтегральные лежат на поверхности и частично погружены в толщу слоя фосфолипидов. Интегральные белки полностью погружены в фосфолипидный бислой и пронизывают его насквозь. Белки выполняют много функций: рецепторную, ферментативную, образуют ионные каналы, являются мембранными насосами и переносчиками ионов и молекул. Полупроницаемость мембран обусловлена несколькими причинами:
Участие мембраны в создании мембранного потенциала. Механизмы активного и пассивного транспорта Рассмотрим трансмембранную разность ионов по обе стороны мембраны (рисунок) K Na Ca Cl над 1 10 20 50 под 50 1 1 1 Разная концентрация ионов по обе стороны мембраны и ведет к электрической асимметрии - разностью зарядов между протоплазмой и поверхностью клетки. Еще Ю. Бернштейн ведущую роль в формировании МП отводил ионам К, считая, что мембрана проницаема только для него. К во много раз больше над мембраной. В целом сумма катионов над мембраной оказывается больше, чем анионов, что и определяет положительный заряд внешней поверхности клеток. В клетке в ходе метаболизма образуется большое количество органических кислот: таких как аспарагиновая, глютаминовая, уксусная, молочная, фумаровая, янтарная. Они диссоциируют с образованием анионов. В протоплазме их содержание значительно превышает количество катионов, а так как они очень крупные, то не могут пройти через поры мембраны и накапливаются у ее внутренней поверхности, чем и обеспечивают отрицательный заряд. Полупроницаемая мембрана участвует в создании ионной асимметрии по обе стороны. Это обеспечивается "активным" и "пассивным" транспортом ионов. В состоянии покоя соотношение констант ионной проницаемости имеет следующий вид: PK : PNа: PCl = 1: 0,04 : 0,45 Пассивный транспорт через мембрану в клетку и из нее в окружающую среду протекает без существенных затрат энергии. Перенос веществ осуществляется по градиенту концентрации. Градиент - это условие, необходимое для переноса веществ через мембрану. Основным механизмом, обеспечивающим проникновение их в клетку, является диффузия. Кроме этого, в формировании мембранного потенциала играют роль силы электростатического взаимодействия. Для лучшего понимания поляризации мембран предположим, что они проницаемы только для ионов К. В клетке К в 50раз больше, поэтому за счет сил диффузии он выходит на поверхность и заряжаете положительно. Анионы орг. кислот, образующих соединения с К из-за большого размера не способны проникнуть через мембрану, поэтому они скапливаются у внутренней поверхности мембраны, образуя ее отрицательный заряд и электростатически удерживают К у внешней поверхности, не давая ему уходить. Таким образом, в мембране возникает электрическое поле, дальнейший выход К задерживается положительным зарядом мембраны и отрицательным зарядом цитоплазмы. В конце концов наступает равновесие Доннана. Оно является одной из главных причин существования МП. Для нейронов при существующей концентрации за счет диффузии К создается мембранный потенциал в 90мв. Но при измерении оказалось, что его величина составляет -70мВ. Это обусловлено тем, что в создании МП имеют значение и ионы Nа. Для него мембрана проницаема намного меньше, чем для К. Но тем не менее за счет диффузии и электростатических взаимодействий Na в небольшом количестве поступает внутрь клетки и снижает МП дол -70мВ. Определенную роль играют и ионы Cl, которых над мембраной в 50 раз больше, чем в цитоплазме. За счет концентрационного градиента они по закону диффузии проникают в клетку, увеличивая ее отрицательный заряд. Однако этому препятствует отрицательный заряд цитоплазмы и положительный заряд мембраны, это приводит к тому, что Cl выталкивается из клетки наружу. Наблюдается равновесие потоков под и на мембрану, которое поддерживается по механизму Доннана. Нужно подчеркнуть, что основная роль в формировании МП принадлежит ионам К, что дало основание для названия теории мембранных потенциалов – калиевой теорией. Натриево-калиевый насос или помпа Однако пассивные механизмы не позволяют понять причины сохранения ионной асимметрии на протяжении всей жизни клетки, кроме того, было замечено, что многие вещества проходят через мембрану против градиента концентрации. Естественно, что этот процесс протекает с затратой энергии. Поэтому, такой механизм переноса называется активным. Активный перенос всегда является избирательным. Он был обнаружен в 1955 году Ходжкиным и назван калий-натриевый насос. Он обеспечивает "откачивание" ионов натрия из клетки и транспорт ионов калия внутрь ее. Осуществляется это с помощью белка-переносчика. Он захватывает в цитоплазме клетки 3 иона натрия и переносят их наружу, где ионы отщепляются и таким образом выводятся из клетки. На наружной поверхности к переносчику присоединяются 2 иона калия, которые закачиваются внутрь клетки. Работа эта осуществляется с затратой энергии, источником которой является аденозинтрифосфат (АТФ). Распад АТФ происходит под действием фермента АТФ-азы, при этом выделяется энергия, которая используется в работе калий-натриевого насоса. При сдвигах трансмембранной концентрации ионов, активность К-Na-насоса может автоматически регулироваться. В регуляции особое значение имеет аденозинтрифосфатаза, которая активируется при увеличении концентрации натрия в цитоплазме и калия в межклеточной жидкости. Работа насоса приводит к следующим результатам: 1) поддерживает высокую концентрацию ионов К+ внутри клетки, обеспечивая тем самым постоянство величины потенциала покоя, 2) поддерживает низкую концентрацию ионов натрия внутри клетки, 3) поддерживая концентрационный градиент натрия, натрий-калиевый насос способствует сопряженному транспорту аминокислот и глюкозы через клеточную мембрану. Таким образом ионная асимметрия обусловлена как избирательной проницаемостью мембраны в состоянии покоя, так и деятельностью К-Na-насоса. Эту величину можно рассчитать по формуле Гольдмана: RT PK [K]Bн + PNа [Nа]Bн + PCl [Cl]H Ем = ______ • ln________________________________________________, где NF PK [K]Bн + PNа [Nа]Bн + PCl [Cl]H PK, PNа, PCl – проницаемость для ионов К, Nа, Cl, вн, н – их внутренняя и наружная концентрация. Изменение мембранного потенциала. Потенциал действия или токи действия Биотоки наблюдаются не только при покое, но и при возбуждении тканей. Электрические процессы всегда сопровождают возбуждение и являются лучшим его критерием. Впервые наличие биотоков при возбуждении было обнаружено Маттеучи в 1837 году в следующем опыте. Он брал 2 н.-м. препарата и нерв одного из них накладывал на мышцу другого, нерв которого раздражался электрическим током. при включении Эл. тока сокращалась не только раздражаемая мышца, но и другая. Этот факт объясняется тем, что при сокращении первой мышцы в ней возникают биотоки, сила которых достаточна для того, чтобы возбудить лежащий на ней нерв второго препарата и вызвать сокращение иннервируемой мышцы. В 1954 году Мюллер и Кёлликер установили, что электрические явления сопровождают и деятельность сердца. Они накладывали на сокращающееся сердце теплокровного животного нерв н.-м. препарата икроножной мышцы лягушки и наблюдали, что при каждом сокращении сердца одновременно сокращается и мышца. Биотоки сердца возбуждают нерв, а он – мышцу. В дальнейшем биотоки были обнаружены во всех возбудимых тканях при их деятельности. В 1800 году Герман назвал токи, сопровождающие процесс возбуждения, потенциалами или токами действия. Этот термин применяется и в наши дни, а токи действия считаются лучшим показателем возбуждения тканей. Токи действия можно зарегистрировать. Это делают микроэлектродным способом. Один электрод располагают на поверхности, а микроэлектрод вводят в клетку. При этом регистрация идет на фоне токов покоя или мембранного потенциала. Сразу после введения электрода внутрь клетки осциллограф регистрирует наличие потенциала покоя, который равен – 70 мв. Если после этого раздражать клетку надпороговым раздражителем, действующим рядом с внеклеточным электродом, то клетка возбуждается и осциллограф записывает кривую однофазного тока действия, которая отражает быстрое колебание мембранного потенциала. В момент возбуждения кривая круто поднимается вверх, доходит до 0 и затем превышает его. После этого возбуждение покидает точку воздействия и заряд мембраны восстанавливается до -70мв. При этом регистрируется однофазный потенциал действия (рис.8). В кривой однофазного тока действия выделяют несколько частей. Восходящую часть кривой называют фазой деполяризации, поскольку она отражает процесс уменьшения и исчезновения исходной поляризации мембраны. Эта фаза протекает наиболее быстро. Вершину тока действия называют спайком. Нисходящее колено характеризует восстановление исходной поляризации мембраны и называют фазой реполяризации. В этой фазе различают 2 части – быстрой реполяризации с крутым падением кривой и медленной, когда восстановление мембранного потенциала замедляется, Эту часть нередко называют следовым отрицательным потенциалом. После него в некоторых тканях (безмякотных нервах) наблюдается следовой положительный потенциал, увеличение заряда мембраны, ее гиперполяризация. Ионный механизм потенциала действия впервые попытался объяснить Ю. Берншетейн в 1912 году с позиции «теории прорыва ионного барьера». Согласно этой гипотезе, при действии раздражителя мембрана теряет свою избирательность и все ионы получают возможность двигаться по своим концентрационным градиентам: Na – в клетку, К – на поверхность. Их концентрация над и под мембраной выравнивается и мембранный потенциал в возбужденном участке исчезает. Это длиться очень короткое время, после чего мембранный потенциал полностью восстанавливается. По Бернштейну амплитуда токов действия равна величине мембранного потенциала. Эта теория была распространена до микроэлектродных исследований Ходжкина и Катца (1949). В своих опытах на гигантских нервных волокнах кальмара ими было установлено, что токи действия имеют большую величину, чем токи покоя: МП при возбуждении не просто падает до 0, а изменяется на противоположный - наружная поверхность заряжается отрицательно по отношению к внутренней. Ходжкиным, Хаксли, Катц (1952) впервые выдвинули теорию об индивидуальном участии различных ионов в формировании потенциала действия (рис.9). Согласно этой теории потенциал действия имеет несколько фаз: 1) фаза градуальной деполяризации – это время от момента нанесения раздражителя до достижения уровня критической деполяризации, после чего развивается высокоамплитудная часть потенциала действия. Градуальная деполяризация характеризуется постепенным раскрытием натриевых каналов, медленным вхождением ионов натрия в клетку по концентрационному градиенту и постепенным снижением МП. Длительность первой фазы для нервной ткани - 0,00004 сек, для скелетной мышцы – 0,0001 сек. При снижении мембранного потенциала до Екр, происходит открытие всех натриевых каналов и развивается следующая фаза. 2) фаза быстрой деполяризации - это время развития пика от начала его возникновения до вершины. Открываются все натриевые каналы, и ионы натрия лавинообразно поступают внутрь клетки по концентрационному и электрохимическому градиенту. В эту фазу смещение мембранного потенциала протекает стремительно, он снижается и приобретает положительный заряд, достигающий величины +30-+40 мВ. Это называется пиком деполяризации или спайком. Амплитуда потенциала действия равна 100-120 мВ. Д 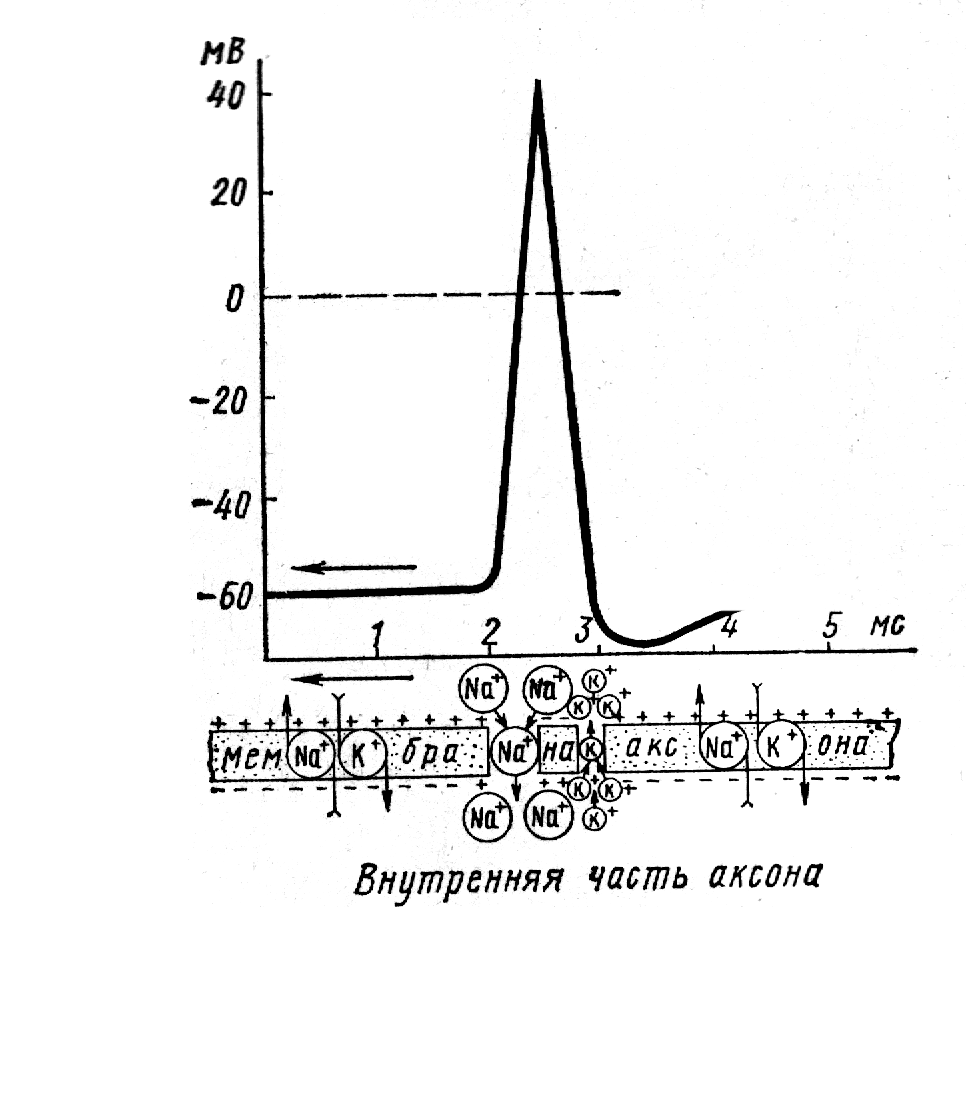 лительность этой фазы для нерва равна приблизительно 0,001-0,002 сек, для мышцы – приблизительно 0,005 сек. лительность этой фазы для нерва равна приблизительно 0,001-0,002 сек, для мышцы – приблизительно 0,005 сек. 3) фаза реполяризации – определяется временем снижения мембранной поляризации до исходного уровня. Начинается в момент достижения заряда мембраны +30-+40мВ. В этот момент инактивируются натриевые каналы и активируются калиевые каналы. Проницаемость для ионов калия увеличивается и он начинает выходить из клетки. Этот период имеет два отрезка времени – относительно быстрое снижение поляризации мембраны (быстрой реполяризации), и последующее более медленное снижение поляризации клетки (медленная реполяризация), которое называется отрицательный следовой потенциал. Медленное снижение мембранной поляризации обусловлено включением в работу активных механизмов переноса ионов натрия и калия (калий-натриевый насос). Длительность третьей фазы для нерва равна 0,02-0,03 сек, для мышцы - приблизительно 0,05-0,1 сек. 4) фаза гиперполяризации (положительный следовой потенциал) – снижение поляризации клеточной мембраны ниже исходной величины. Гиперполяризация характерна для немиелинизированных нервных волокон. Ее связывают с временно увеличенной проницаемостью для ионов К+. Длительность следовой электроположительности для нерва приблизительно равна 0,1 сек, для мышцы – 0,25 сек и больше. После гиперполяризации МП полностью нормализуется до исходных -70мВ. Подобные ПД наблюдаются в любой возбудимой системе, протекая с различной скоростью и занимая различное время. ПД развивается по закону «все или ничего». Токи действия служат одним из самых объективных критериев возбуждения, поэтому их регистрация используется для оценки работы многих органов: ЭКГ, ЭЭГ, электромиография и т.д. Токи действия нашли практическое применение в протезировании – в создании управляемых протезов. |