Лекция 1. Микробиология в пищевой промышленности. Проблемы систематики в микробиологии. Вопросы
 Скачать 0.52 Mb. Скачать 0.52 Mb.
|

|
Генетика микроорганизмов Вопросы: 1.Наследственные факторы микроорганизмов. 2.Механизмы, вызывающие изменение генетической информации. 3.Практическое использование достижений генетики микроорганизмов 1. Наследственные факторы микроорганизмов. В клетках эукариот местом нахождения генетического материала являются ядра, а у прокариот – нуклеоиды. Генетический материал представлен ДНК. Бактериальные клетки ДНК имеют форму нитей, замкнутых в виде кольца, - бактериальная хромосома. Хромосома имеет отдельные участки (фрагменты молекулы ДНК), которые называются генами. Ген – основной фактор, отвечающий за наследственные свойства микроорганизмов. Кроме того, конкретные признаки микроорганизмов обуславливают отдельные ферменты. Гены, которые несут информацию о синтезируемых микроорганизмами ферментах – структурные гены. Микроорганизмы содержат генетический материал не только в хромосоме, но и в плазмидах, расположенных в цитоплазме. Плазмиды представляют собой молекулы ДНК. Клетка микроорганизма составляет генотип данного микроорганизма. Проявление наследуемых морфологических признаков и физиологических процессов называется фенотипом. Изменения наследственных признаков, возникающие под влиянием внешней среды, - модификации. Модификации существуют до тех пор, пока действует вызывающий их фактор среды, и не наследуются организмами. Изменения генотипа называются мутациями, они происходят случайно и являются наследственно закрепленными признаками. 2. Механизмы, вызывающие изменения генетической информации. Мутации происходят, если в ДНК химически изменяется или выпадает нуклеотид или в ДНК включается лишний нуклеотид. Различают генные и хромосомные мутации. Генные мутации затрагивают только 1 ген, а хромосомные распространяются на несколько генов. Генные мутации:
В ряде случаев точковые мутации могут возвращаться к исходной дикой форме в результате процесса обратной мутации – реверсии. Хромосомные мутации связаны с более крупными перестройками фрагментов ДНК. Среди них выделяются:
Мутации вызывают обычно химические и физические агенты, такие как рентгеновское, ультрафиолетовое излучения, гамма-лучи, соединения тяжелых металлов, перекиси, минеральные масла, алкилирующие соединения, аналоги иприта и другие. Клетки бактерий обладают специальными системами, восстанавливающими поврежденные ДНК. Восстановления осуществляются ферментами, которые находятся под контролем специальных генов. У микроорганизмов имеются механизмы, способствующие возникновению в потомстве резко измененнной наследственности. Эти механизмы заключаются в немедленной перестановке генов (рекомбинации), принадлежащих близкородственным, но генетически различным организмам. У эукариот это образование индивидуумов происходит в результате полового процесса. У прокариот известно 3 процесса рекомбинации генов:
3. Практическое использование достижений генетики микроорганизмов. Развитие генетики, открывшей методы получения наследственно измененных форм микроорганизмов, расширило возможности использования микроорганизмов в сельскохозяйственном и промышленном производстве, а также в медицине. Основной из этих методов — это индуцированное получение мутантов воздействием различными мутагенами (излучениями и химическими веществами) на дикие, существующие в природе культуры микроорганизмов. Таким методом удается создать мутанты, которые дают в десятки и сотни раз большее количество ценных продуктов (антибиотиков, ферментов, витаминов, аминокислот и т. д.) по сравнению с дикими формами микроорганизмов. Процесс получения высокопродуктивных штаммов микроорганизмов состоит из многих этапов. На культуру микроорганизма воздействуют различными мутагенными факторами с последующим отбором наиболее продуктивного штамма. Этот мутантный штамм может подвергнуться дальнейшему воздействию мутагенов и дальнейшему отбору еще более продуктивных мутантов. Часто из тысячи бесполезных мутантов отбирают только один высокопродуктивный штамм. В последние годы методом радиационного и химического мутагенеза микроорганизмов получено большое число промышленных штаммов микроорганизмов — продуцентов антибиотиков, ферментов, витаминов, ценных пищевых аминокислот, ростовых и других веществ. Особенно широкие перспективы переделки наследственной природы организмов сулит развитие генной, или генетической, инженерии. Это раздел молекулярной генетики, который разрабатывает методы создания новых генетических структур, несущих заданную информацию, и способов их переноса в клетки прокариот и эукариот. Полученные методом генной инженерии новые генетические молекулы представляют собой рекомбинантные ДНК, включающие два компонента — вектор (переносчик) и клонируемую «чужеродную» ДНК. Так как переносчик должен обладать свойствами репликона и обусловливать репликацию вновь созданной рекомбинантной ДНК, то в качестве вектора обычно используют такие репликоны, как плазмиды, умеренные фаги и вирусы животных. Все эти переносчики имеют циркулярно замкнутую структуру ДНК - Клонируемая ДНК — это фрагмент ДНК, который несет необходимый ген (или гены), контролирующий образование нужного вещества. Имеются различные приемы получения рекомбинантных молекул ДНК - Наиболее простой из них сводится к обработке изолированных молекул ДНК-вектора и ДНК, несущей необходимый ген, ферментами рестриктазами (эндонуклеазы рестрикции), расщепляющими взятые молекулы ДНК в строго определенном месте с образованием однонитчатых комплементарных друг другу концов, так называемых липких концов. Это первый этап получения рекомбинантных ДНК — «разрезание» молекул ДНК с помощью эндонуклеаз рестрикции. Второй этап заключается в обработке полученных линейных молекул ДНК ферментом полинуклеотидлигазой, которая «сшивает» две разные молекулы в одну рекомби-нантную ДНК. На третьем этапе рекомбинантные молекулы вводят в клетки тех или иных бактерий методом трансформации. На завершающем, четвертом, этапе проводят клонирование трансформированных клеток. В настоящее время методом генной инженерии получены рекомбинантные молекулы ДНК, несущие информацию для образования таких важных веществ, как интерферон, инсулин, гормон роста человека и другие в клетках кишечной палочки (Е. coli). По-видимому, методом генной инженерии можно будет создать и такие бактерии, которые, потеряв свою болезнетворность, помогут выработать иммунитет против многих инфекционных болезней животных и человека. В промышленности, благодаря использованию генной инженерии, появятся высокопродуктивные микроорганизмы, создающие белки, ферменты, витамины, антибиотики, ростовые вещества и другие нужные продукты. Будут получены новые сорта растений и породы животных, устойчивые к заболеваниям и наделенные особенно выгодными для сельского хозяйства свойствами. Возможно, методом генной инженерии будут созданы растения, обладающие способностью к связыванию молекулярного азота атмосферы. Такие растения, вероятно, можно будет получить после введения в их геном генов от микроорганизмов, фиксирующих азот из воздуха. Нет сомнения в том, что в связи с разработкой и совершенствованием методов генной инженерии, показавших возможность передачи не только естественных генов живых организмов, но и искусственно синтезированных, открываются блестящие перспективы для научно-технического прогресса не только в медицине и промышленности, но и в сельскохозяйственном производстве. ЛЕКЦИЯ №6 Превращение органических веществ, не содержащих азот. Вопросы: 1.Спиртовое и молочнокислое брожения. пропионовокислое, маслянокислое брожения. 2.Ацетоно-бутиловое, ацетоно-этиловое брожения, брожение клетчатки и пектиновых веществ. 3.Процессы окисления. У микроорганизмов различают 2 основные формы катаболизма: брожение и аэробное дыхание. Все эти процессы происходят под действием ферментов микроорганизмов. Причем у микроорганизмов различают внутриклеточные ферменты (эндоферменты), которые участвуют в метаболизме внутри клетки и содержатся в ней; и внеклеточные ферменты (экзоферменты), которые выделяются клетками микроорганизмов в окружающую среду. Экзоферменты в основном относятся к классу гидролаз и разлагают соединения большой молекулярной массы, которые не могут проникнуть в клетку микроорганизма. Продукты разложения легко проникают в клетку и используются в качестве питательных веществ. В разнообразии ферментов, выделяемых в окружающую среду, и заключается роль микрофлоры в круговороте веществ в природе.
Брожение - анаэробное разложение углеводов на продукты, которые далее не разлагаются без участия молекулярного кислорода. У различных микроорганизмов продукты разложения различны и зависят от набора ферментов и внешних условий. Спиртовое брожение. Основной возбудитель спиртового брожения – дрожжи, применяемые в хлебопечении, при получении спирта, вина, пива, кваса и некоторых молочнокислых продуктов (кефир, кумыс). В бродильных производствах используют представителей родов Saccharomyces и Scizosaccharomyces. В небольших количествах спирт может накапливаться в средах, богатых углеводами, при развитии в них некоторых видов грибов родов Aspergillus, Mucor, Fusarium, а также бактерий родов Sarcina и Zimomonas. При доступе кислорода дрожжи, вызывающие брожение, начинают окислять углеводы, то есть переходят к аэробному дыханию с образованием углекислого газа и воды. Аэробное дыхание дает больше энергии, чем брожение. Коэффициент использования углеводов повышается и поэтому при производстве дрожжей для получения их большей массы среду, в которой происходит их размножение, аэрируют. Сбраживание углеводов дрожжами с образованием этилового спирта и углекислого газа идет по гликолитическому пути (путь Эмбдона – Мейергофа - Парнаса). Превращение происходит в результате сложных ферментативных реакций, осуществляемых через ряд промежуточных стадий.  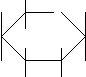 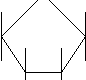 СН2ОН СН2ОР О СН2ОН СН2ОР О    Н О Н Н О Н СН2ОР СН2ОН Н + АТФ Н изомераза Н О Н Н О Н СН2ОР СН2ОН Н + АТФ Н изомераза   ОН Н ОН Н Н ОН ОН Н ОН Н Н ОН ОН ОН ОН ОН Н ОН ОН ОН ОН ОН Н ОНН ОН Н ОН ОН Н глюкозо–6–фосфат фруктозо-6-фосфат  О О  СН2ОР СН2ОР СН2ОР СН2ОР СН2ОР СН2ОР СН2ОР СН2ОР+ АТФ Н ОН альдалаза НСОН + С=ОН ОН ОН Н С=О СН2ОН Н фруктозо-1,6-дифосфат триозы Эти триозы под действием триозофосфатизомеразы легко переходят друг в друга, и дальнейшему превращению подвергается фосфоглицериновый альдегид.  СН2ОР СН2ОР СН2ОР СН2ОН СН2ОР СН2ОР СН2ОР СН2ОН НS-фермент + АДФ изомераза НСОН + АТФ СНОН СНОН СН2ОР НС=О РОС=О НОС=О НОС=О 1,3-дифосфоглицериновая 3-фосфоглицериновая 2-фосфоглицериновая кислота кислота кислота  енолаза + АДФ фосфотрансфераза СН3 СН2=СОР СН2=СОН - Н2О С=О СООН СООН СООН фосфоенолпируват енолпируват пируват декарбоксилаза НАД Н2 СН3СОН СН3СН2ОН - СО2 ацетальдегид этанол Образовавшийся пируват под действием карбоксилазы превращается в уксусный альдегид, выделяется углекислый газ. Затем под действием алкогольдегидрогеназы (НАД Н2) образуется этиловый спирт. В результате спиртового брожения в качестве побочных продуктов может образоваться глицерин, который идет на образование уксусной, молочной, янтарной, пропионовой кислот, диацетила, различных альдегидов и сложных эфиров. Кроме вторичных продуктов образуются побочные продукты: высшие спирты, называемые сивушными маслами (изоамиловый, бутиловый, пропиловый спирты и некоторые ароматические спирты). Эти вещества образуются из соответствующих кетокислот, синтезирующихся в процессе метаболизма углеводов. Спиртовое брожение протекает при кислой реакции среды рН 4-5. В щелочных условиях рН 8 один из основных продуктов брожения – глицерин. Наибольшее практическое значение в спиртовом брожении имеет вид дрожжей Saccharomyces cerevisiae. Дрожжи сбраживают не все сахара, а чаще всего гексозы; пентозы сбраживает ограниченное число видов. Дисахариды также сбраживают некоторые виды микроорганизмов, причем должен быть строго определенный набор сахаров. Перед сбраживанием полисахариды под действием ферментов дрожжей распадаются на моносахариды. Температура, при которой дрожжи могут развиваться, колеблется от 0оС до 50оС, но оптимальная температура для роста большинства видов 28 – 30оС. В микробиологии виды делят на штаммы, а штаммы на расы. Среди штаммов различают расы верхового и низового брожения. Дрожжи верхового брожения используют для брожения при 14 – 25оС, в этих условиях обильно выделяется углекислый газ, наблюдается пенообразование, клетки микроорганизмов поднимаются на поверхность жидкости. Эти дрожжи используют в хлебопечении, спиртовой промышленности. Дрожжи низового брожения применяют в производстве при 0 – 10оС, брожение происходит спокойно, и клетки остаются на дне сосуда. Такие расы используют в пивоварении и виноделии (Saccharomyces vini). В промышленности используют чистые культуры дрожжей, которые обеспечивают наиболее правильное течение процесса и получение качественной продукции. Среди дрожжей имеются вредные для человека виды, некоторые из них способны вызывать окисление углеводов, есть вредители пищевой продукции и вин. Молочнокислое брожение. Это брожение вызывает специфическая группа молочнокислых бактерий, в результате происходит распад глюкозы до молочной кислоты. Известно 3 типа брожений, вызываемых молочнокислыми бактериями.
С  6Н12О6 2 СН3СНОНСООН 6Н12О6 2 СН3СНОНСООН Лактат В основе гомоферментативного брожения лежат реакции гликолиза, образующийся пируват восстанавливается до лактата водородом под действием фермента лактатдегидрогеназы. 2 НАД Н2 С  Н3СОСООН 2 СН3СНОНСООН Н3СОСООН 2 СН3СНОНСООН
С  6Н12О6 СН3СНОНСООН + С2Н5ОН + 2 СО2 6Н12О6 СН3СНОНСООН + С2Н5ОН + 2 СО2лактат У гетероферментативных молочнокислых бактерий отсутствуют главные ферменты гликолиза, такие как триозофосфатизомераза и фруктозодифосфатальдолаза. Поэтому начальное превращение глюкозы идет по пентозофосфатному пути до образования рибулозо-5-фосфата, который затем превращается в ксилулозо-5-фосфат, а тот расщепляется с образованием фосфоглицеринового альдегида и ацетилфосфата. Фосфоглицериновый альдегид превращается в пируват, затем в лактат, как в процессе гликолиза. Из ацетилфосфата образуется этиловый спирт и уксусная кислота.
2 С6Н12О6 3 СН3СООН + 2 СН3СНОНСООН ацетат лактат Бифидоброжение осуществляется по пентозофосфатному пути с конечными продуктами в виде уксусной кислоты и лактата. Пентозофосфатный путь известен как путь Энтнера – Дудорова. Иногда в сбраживаемых средах накапливается небольшое количество диацетила и ацетоина (вещества, обладающие приятным ароматом, который передается продукту). Оптимальными параметрами для роста молочнокислых бактерий являются рН 5,5-8,8. Палочковидные формы развиваются в более низких значениях рН, чем кокковидные. Благоприятная температура от 7оС до 42оС, но температурный оптимум роста от 30оС до 40оС. молочнокислые бактерии не образуют спор, поэтому при температуре выше предела погибают. Кроме глюкозы молочнокислые бактерии сбраживают фруктозу, лактозу, сахарозу, мальтозу и некоторые пентозы. Молочнокислые бактерии могут быть 2-х видов: молочнокислые стрептококки и молочнокислые палочки. Молочнокислые бактерии применяются при изготовлении сырокопченых колбас, в квашении овощей. Пропионовокислое брожение. Возбудителями этого вида брожения являются пропионовокислые бактерии, представляющие собой палочки, расположенные по одиночке, парами, или цепочками. Пропионовокислые бактерии являются анаэробами. Продуктами брожения углеводов могут быть различные комбинации пропионовой и уксусной кислот и небольшие количества изовалериановой, муравьиной, янтарной кислот или молочной кислоты и углекислого газа. Сбраживание углеводов пропионовокислыми бактериями происходит по гликолитическому пути до образования пирувата, в зависимости от условий пируват может окисляться, карбоксилироваться или восстанавливаться. Суммарное уравнение: 3  С6Н12О6 4 СН3СН2СООН + СН3СООН + 2 СО2 + 2 Н2О С6Н12О6 4 СН3СН2СООН + СН3СООН + 2 СО2 + 2 Н2Оглюкоза пропионовая уксусная кислота кислота 3 СН3СНОНСООН 2 СН3СН2СООН + СН3СООН + СО2 + Н2О лактат Для роста пропионовокислых бактерий температурный оптимум 30 – 37оС, реакция среды рН = 7. Пропионовокислые бактерии обнаруживаются в пищеварительном тракте жвачных животных, их используют в производстве твердых сычужных сыров. По окончании молочнокислого брожения лактозы в сыре наступает стадия пропионовокислого брожения, которое сопровождается сбраживанием молочной кислоты и других веществ. Углекислый газ обуславливает появление рисунка сыра (глазки). Процессы брожения, вызываемые бактериями рода Clostridium и Enterobacterium. Бактерии рода Clostridium могут сбраживать те или иные органические соединения, в зависимости от вида их делят на несколько групп.
М  аслянокислое брожение. Источником углерода для маслянокислых бактерий служат моносахариды, дисахариды, крахмал, декстрины, молочная кислота, пируват, глицерин. Маслянокислое брожение начинается со сбраживания сахара по гликолитическому пути, затем при участии ферментов синтезируется масляная кислота; как побочный продукт – уксусная кислота, происходит восстановление некоторых соединений. В результате брожения образуется в основном углекислый газ, водород, масляная и уксусная кислоты. аслянокислое брожение. Источником углерода для маслянокислых бактерий служат моносахариды, дисахариды, крахмал, декстрины, молочная кислота, пируват, глицерин. Маслянокислое брожение начинается со сбраживания сахара по гликолитическому пути, затем при участии ферментов синтезируется масляная кислота; как побочный продукт – уксусная кислота, происходит восстановление некоторых соединений. В результате брожения образуется в основном углекислый газ, водород, масляная и уксусная кислоты.4 С6Н12О6 3 СН3СН2СН2СООН + 2 СН3СООН + 8 СО2 + 8 Н2 глюкоза масляная кислота уксусная кислота Маслянокислые бактерии являются строгими анаэробами, среди них есть патогенные (Clostridium botulinum, Clostridium perfringers), сапрофитные формы. Маслянокислое брожение не всегда желательный процесс, например, в квашеных продуктах при развитии маслянокислых бактерий разлагается белковая часть, масляная кислота придает продуктам неприятный запах.
Бактерии рода Enterobacterium (энтеробактерии) – представители кишечной микрофлоры, могут осуществлять смешанное – кислое и бутандиоловое – брожение, при котором образуются некоторые органические кислоты, спирты, углекислый газ и водород. Эти бактерии являются факультативными анаэробами. Превращение углеводов у них происходит по гликолитическому пути. К энтеробактериям относятся следующие роды:
К энтеробактериям также относятся Proteus, Enterobacter и другие организации. Брожение клетчатки и пектиновых веществ. Брожение клетчатки заключается в разрушении клетчатки в анаэробных условиях под влиянием целлюлазы до целлобиозы, которая затем переходит в глюкозу, а глюкоза сбраживается с образованием масляной, уксусной кислот, углекислого газа, водорода, метана. Один из типичных представителей, сбраживающих клетчатку, - это бактерии Омелянского. Брожение пектиновых веществ протекает под действием протопектиназы, пектиназы, пектазы. Сбраживание происходит с образованием уксусной кислоты, этилового спирта, углекислого газа, водорода. |