+3-лек Поступл ВОДЫ в клетку. Лекция 3 поступление воды в растительную клетку
 Скачать 4.48 Mb. Скачать 4.48 Mb.
|

|
ЛЕКЦИЯ 3 ПОСТУПЛЕНИЕ ВОДЫ В РАСТИТЕЛЬНУЮ КЛЕТКУ План Структура и свойства воды Поступление воды в растительную клетку (классическая теория). Методы определения осмотического давления и сосущей силы клеток Поступление воды в клетку с термодинамических позиций Влияние различных условий на поступление воды в клетку и водный потенциал клетки Структура и свойства воды Вспомните из неорганической химии структуру молекулы воды. Общая электронная пара смещена к кислороду, как более электроотрицательному атому, что обеспечивает полярность молекулы, дипольность. Свойства воды Растворимость. Эта структурная особенность обусловливает важнейшее свойство воды - растворять вещества лучше, чем другие жидкости. Растворение кристаллов неорганических солей осуществляется благодаря гидратации входящих в их состав ионов. 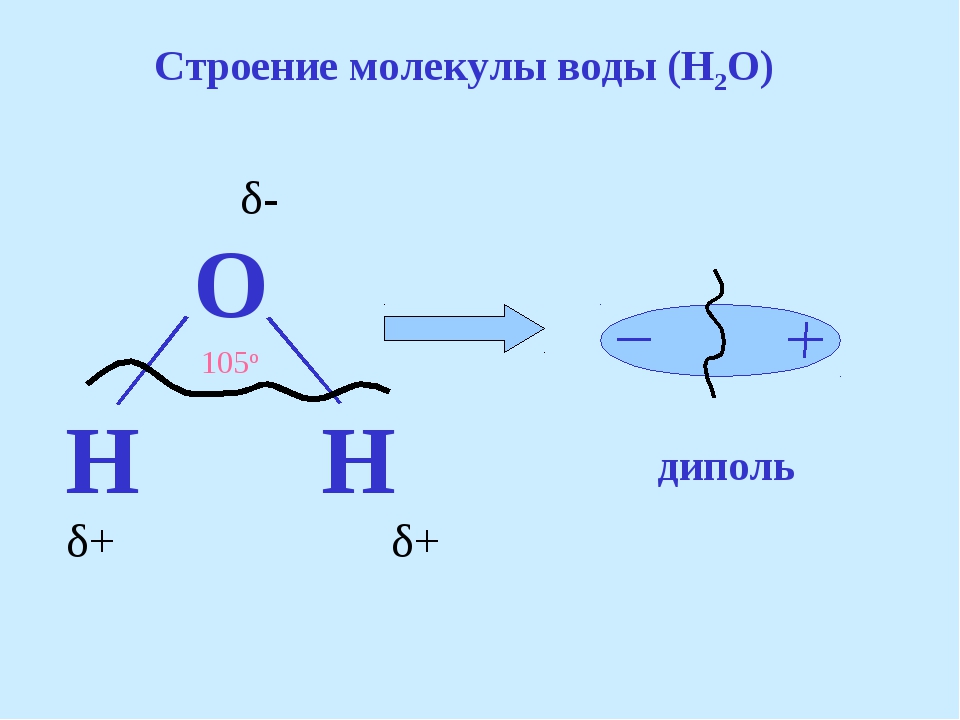   Высокая теплоемкость. Молекулы воды соединены между собой водородными связями. В твердом состоянии все молекулы соединены между собой. При 0° - С 15% водородных связей разрывается, при 20°С - 20%. Соединенные между собой молекулы образуют кластеры («гроздь, пучок»). На разрыв водородных связей требуется затрата энергии. Благодаря этому вода обладает высокой теплоемкостью - количество теплоты, необходимое для повышения температуры на 1° С. Теплоемкость воды в 5 - 30 раз выше, чем у других веществ. Например, при одинаковом нагреве солнцем вода в водоеме нагревается в 5 раз слабее, чем песок на берегу, но во столько же раз дольше сохраняет теплоту. Когезия. Благодаря силам сцепления молекул воды - когезия - на поверхности воды создается поверхностное натяжение (выше - только у ртути). (Док-ва когезии: по поверхности воды перемещаются водомерки, мелкие ящерицы..; в полном стакане воды создается «горочка» (как и в сосудах и микрокапиллярах между фибриллами целлюлозы растений – играет роль в усилении и ослаблении скорости транспирации) Адгезия. Благодаря адгезии - прилипанию - вода поднимается против гравитационных сил по проводящей системе. Функции воды в клетке. 1. Универсальный растворитель. Создает необходимые условия для реакций обмена веществ. 2. Транспортная. Обеспечивает перемещение веществ по проводящей системе растения. 3. Вода является реагентом т.к. участвует в целом ряде химических реакций именно как химическое вещество: фотосинтез, гидролиз веществ. 4. Упорядочение структур в клетке. Макромолекулы приобретают определенную конформацию благодаря гидратным оболочкам. 5. Обеспечивает упругое состояние клеток (состояние тургора). 6. Терморегуляция и термостабилизация. Высокая теплоемкость воды защищает растения от резкого повышения температуры во время жары, а высокая теплота парообразования способствует охлаждению. В клетке не возникает «горячих точек» от экзотермических реакций благодаря высокой теплопроводности. (Пример: выходя из реки в жаркую погоду испытываем холод). 7. Объединяет все молекулы и структуры в клетке в единое целое. Формы воды в клетке. С химических позиций в растительной клетке присутствует две формы воды: • свободная - участвует в химических реакциях, испаряется, транспортируется. • связанная - гидратирует молекулы и ионы. В зависимости от того, какие частицы гидратируются различают: - коллоидносвязанную воду (гидратирует плохо растворимые, крупные органические молекулы, например, белки). -осмотическисвязанную воду (гидратирует растворенные вещества - молекулы и ионы, например, соли, кислоты, сахара). Физиологическое значение этих форм воды различно. Интенсивность физиологических процессов зависит от содержания свободной воды. В молодых клетках ее содержится больше. Устойчивость клеток к действию неблагоприятных факторов зависит от содержания связанной воды, что характерно для старых клеток. Распределение воды в клетке. Содержание воды в различных структурах клетки неодинаково: - Наиболее богата водой вакуоль (до 98%). Здесь содержится свободная и осмотически связанная вода. - Наименьшее содержание воды в клеточной стенке (= 40%), где находится свободная и коллоидносвязанная вода, т. к. там присутствуют высокополимерные молекулы. - В цитоплазме содержатся все формы воды (до 95%). В целом содержание воды в клетке изменяется в зависимости от ее возраста, органа, экологической принадлежности растения, условий существования. Молекулы воды преодолевают биологические мембраны 2-мя путями - через липидный бислой - через поры, или водные каналы, образуемые специфическими белками – аквапоринами. Они были обнаружены в конце 80-х годов20 столетия. Молекула такого белка состоит из 6 субъединиц – а-спиралей, пересекающих 6 раз липидный бислой. Между 2и3-е1, а также между 5 и 6-й спиралями имеются гидрофобные петли – NPA-бокс, содержащий иминокислотные остатки аспарагиновой кислоты, пролина и аланина. Эти петли совмещаются в центре липидного слоя и формируют 2 полусферические поры, которые вместе создают узкий водный канал. Через него легко «проскакивают» молекулы воды (более 1 млн. молекул в секунду в обоих направлениях в зависимости от градиента водного потенциала). Эти белки найдены в клетках практически всех растений и отличаются большим разнообразием. Выделяют 2 группы аквапорин по месту их локализации в клетке: аквапорины плазмаллемы и аквапорины тонопласта. помимо транспорта воды они могут транспортировать маленькие молекулы органических веществ, обеспечивающих осморегуляцию в цитоплазме. 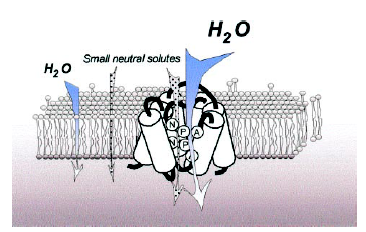 Аквапорины входят в семейство интегральных белков Аквапорины входят в семейство интегральных белков Способы поступления воды в растительную клетку Поглощение воды из внешней среды - обязательное условие существования любой живой клетки. В растительную клетку вода может поступать различными способами. 1. Благодаря набуханию биоколлоидов, увеличению степени их гидратации. Движение воды идет по градиенту концентрации и в клетке развивается давление набухания. Такое поступление воды характерно для сухих семян, помещенных в воду или во влажную почву. Обычно семена при этом набухают, увеличиваются в объеме, поскольку клеточные оболочки. протопласт, запасные вещества представляют высохшие коллоидные мицеллы. Наибольшее набухание наблюдается у белковых веществ, меньше у крахмала, еще меньшее - у клетчатки. белковые вещества > крахмал > клетчатка Поэтому сильнее набухают семена богатые белками (горох, фасоль). Значительно меньше набухают семена злаков. Набухание семян приводит к разрыву семенной кожуры, состоящей из менее разбухающих веществ, чем внутреннее содержимое. В совершенно сухих семенах может развивать давление около 1000 атмосфер. В рассказе К. Паустовского «Черное море» описан случай, когда корабль, перевозивший в трюме сухие семена гороха, разломился пополам, получив небольшую пробоину. 2. Активное поступление воды против градиента концентрации, связанное с затратой энергии. Основанием к предположению о существовании активного поступления послужили опыты англ. ученого Беннет-Кларка. При помещении клеток в раствор, концентрация которого была выше концентрации клеточного сока, плазмолиз не наблюдался. Было установлено, что протоплазма активно секретировала определенное количество воды в вакуоль. Поступление воды в клетку уменьшается при недостатке кислорода или введении дыхательных ядов. Это тоже может служить доказательством активного поступления воды в клетку. 3. В некоторых случаях может происходить заглатывание воды клеткой в процессе пиноцитоза. 4. Осмотическое поглощение - основной способ поступления воды в растительную клетку. Для того, чтобы нам разобраться в процессах, происходящих при поступлении воды в клетку, необходимо вспомнить из физики такие понятия, как диффузия и осмос. Диффузия - движение молекул и ионов из области с высокой концентрацией в область с более низкой концентрацией, или иными словами - движение по градиенту концентрации, что приводит к равномерному распределению молекул растворителя и растворенного вещества. Например, чай сахаром, который не размешали ложечкой. Осмос - это переход молекул растворителя (воды) из области с его более высокой концентрацией в область с более низкой через мембрану (полупроницаемую или избирательно проницаемую). Т.е. осмос - это разновидность диффузии, которая осуществляется через мембрану.  Природу осмоса изучали с помощью простой конструкции осмометра. Первый осмометр был сконструирован в 1826 г. фр. физиологом Дютроше: Мешочек из полупроницаемой пленки (пергамент, животный пузырь) с помещенной в него стеклянной трубкой, заполнялся раствором сахарозы. После погружения мешочка в чистую воду наблюдался подъем уровня жидкости в трубке. Однако применявшиеся Дютроше пленки не были абсолютно полупроницаемыми и медленно проникавший сахар мешал количественному измерению осмотического давления. 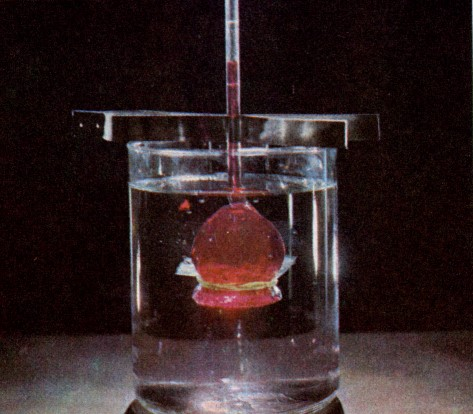 Более совершенная конструкция осмометра была предложена в 1877 г. нем. физиологом Пфеффером. Свой осмометр он назвал «искусственной клеткой». Основой ее служил пористый фарфоровый сосуд. В сосуд наливался раствор медного купороса - CuS04, а затем он погружался в раствор желтой, кровяной соли - К4 [Fе(СN)6]. При взаимодействии этих веществ в порах сосуда образовывалась пленка из гексацианоферрата меди – Сu2[Fе(СN)б], служившая полупроницаемой мембраной (плазмалемма). Раствор медного купороса выливался, и стакан наполнялся раствором сахарозы, который затем помещался в чистую воду. Присоединяя к сосуду манометр, измеряли силу давления, с которой вода поступала в раствор. В системе: раствор - мембрана - растворитель, в результате движения молекул возникает осмотическое давление. Осмотическое давление - это диффузионное давление частиц растворителя, направленное из чистого растворителя в сторону раствора, отделенного полупроницаемой мембраной. Вант-Гофф показал, что осмотические законы соответствуют газовым законам Бойля-Мариотта. Для расчета осмотического давления он предложил формулу: Р = RTci , где С - концентрация в молях, Т - абсолютная температур а (273 +t °C), R -универсальная газовая постоянная (0,082), i - изотонический коэффициент, равный 1+а, где а - степень электролитической диссоциации (отсюда следует, что для неэлектролитов - сахароза - i =1, для электролитов - соли - больше 1 (а см в табл). Осмотическое давление может выражаться в атмосферах, барах, Паскалях. Осмотическое давление связано с тургорным давлением и сосущей силой клетки Силу, с которой клетка поглощает воду, называют сосущей силой - S. Величина сосущей силы определяется осмотическим давлением клеточного сока и тургорным давлениемв клетке - Т. S = Р - Т Тургорное давление - противодавление клеточной стенки, возникающее при ее эластическом растяжении.  Соотношение между величинами S, Р, Т при различных условиях водоснабжения клетки: 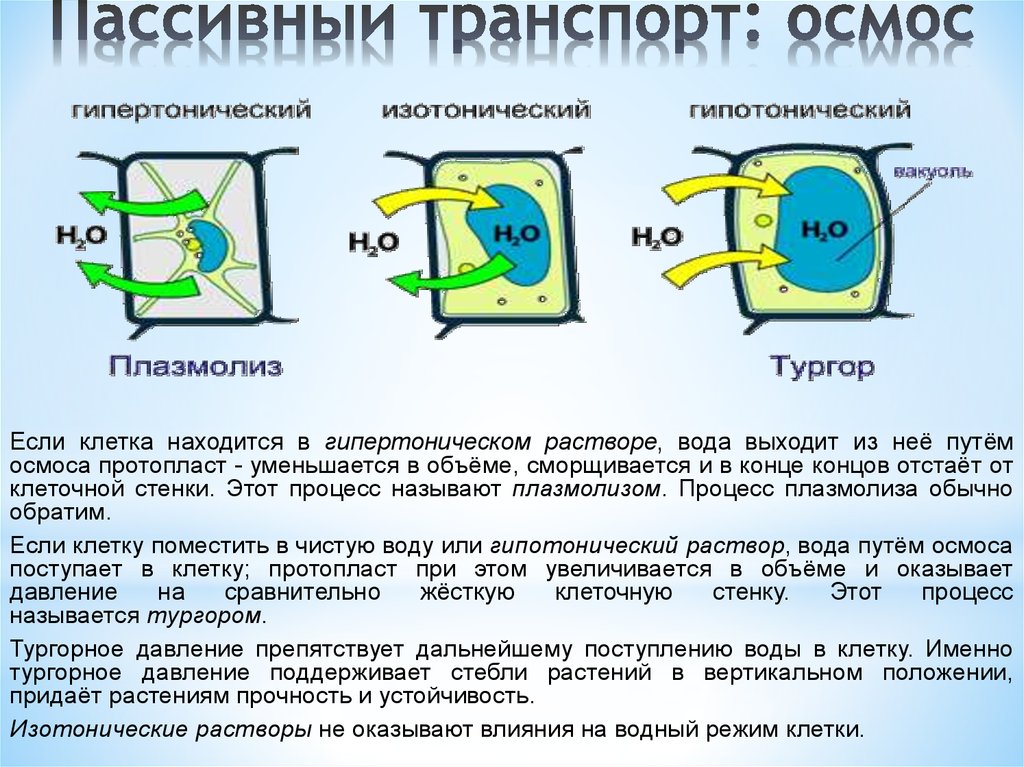 В условиях достаточного насыщения водой: S = Р - Т В состоянии тургора - полного насыщения водой - сосущая сила = 0, а тургорное давление максимально и равно осмотическому (Т-мах): S = 0, Р = Т max . При потере клеткой воды (плазмолиз, завядание клетки) тургорное давление уменьшается и может стать Т = 0, а сосущая сила достигает своего максимального значения и равняется осмотическому давлению (S-max): Т = 0, S max = P. Плазмолиз – отхождение протопласта от клеточной стенки в результате потери клеткой воды при ее погружении в гипертонический раствор Различают: • гипертонический раствор - концентрация веществ больше, чем концентрация веществ в клетке. • гипотонический раствор - концентрация веществ меньше, чем концентрация веществ в клетке. • изотонический раствор - концентрации веществ в клетке и окружающем растворе равны.   
В гипертоническом растворе происходит явление плазмолиза. Пространство между сократившимся протопластом и клеточной стенкой заполняет наружный раствор. Плазмолизируются только живые неповрежденные клетки. Этапы плазмолиза: уголковый вогнутый выпуклый При помещении плазмолизированной клетки в воду или гипотонический раствор наблюдается деплазмолиз. Если гипертонический раствор содержит ион К+, наблюдается особая форма плазмолиза - колпачковый, доказывающий неодинаковую избирательную проницаемость плазмалеммы и тонопласта. Ион К+ легко проникает через плазмалемму и плохо через тонопласт. Накапливаясь в цитоплазме, ионы калия уменьшают ее вязкость, увеличивают объем.. У молодых клеток при потере воды в воздушной атмосфере, например, при суховее, наблюдается явление циторриза. Тургорное давление в клетках становится отрицательным и протопласты, сокращаясь в объеме, не отделяются от клеточных стенок, а тянут и  х вслед за собой. Внешне это проявляется в образовании на поверхности клеток волнообразных изгибов. х вслед за собой. Внешне это проявляется в образовании на поверхности клеток волнообразных изгибов.Сосущая сила таких клеток равна сумме осмотического и тургорного давлений: S = Р - (-Т) = Р + Т . Влияние различных условий на поступление воды в клетку (по классической теории) Величина осмотического давления зависит как от внешних, так и от внутренних факторов. Внешние условия: • Температура • Содержание воды • Концентрация окружающего раствора. Внутренние факторы: 1) Возраст (в молодых клетках - ниже) 2) Экологическая принадлежность (увеличивается в направлении : гидрофиты (1-3 атм) мезофиты (5-10 атм) ксерофиты (20 атм), галофиты (до 60 - 80 атм). Огромные величины осмотического давления в клетках галофитов кажутся невероятными и возникает вопрос, как могут клеточные оболочки выдерживать такое напряжение. Однако раствор засоленных почв, играющий роль окружающей среды, сам обладает высокой концентрацией. Клеточная оболочка в этом случае испытывает на себе воздействие только перевеса внутреннего давления над наружным. Если растение быстро перенести в чистую воду, то клеточные оболочки лопаются. 3) Жизненная форма растения (увеличивается в направлении: травы - кустарники —> деревья). 4) Место расположения клеток по вертикальной оси растения увеличивается в направлении: клетки корня -> стебля —> листьев). Методы определения осмотического давления и сосущей силы клеток. Определение величины осмотического давления имеет больше значение для экологических исследований. Величина осмотического давления позволяет судить о способности растений поглощать воду из почв и удерживать ее. Существует несколько методов определения Р: прямые методы: 1. криоскопический - определяют точку замерзания выжатого сока растения, делают специальный пересчет для определения его концентрации. 2. рефрактометрический - прямое определение концентрации клеточного сока с помощью рефрактометра. косвенные методы: 3. плазмолитический - разработан Де-Фризом (1884). Растительные клетки помещаются в серию растворов известной концентрации, находится изотонический р-р. Для изотонического раствора Sp-pa = Sкл.; Sp-pa = Рр-ра = RTci. 4. метод струек - основан на учете изменения концентрации окружающего раствора при погружении клеток в серию растворов. 5. упрощенный метод по Уршпрунгу - основан на учете изменении размеров растительной ткани при её погружении в серию растворов. Рр-ра = RTci. Поступление воды в клетку с термодинамических позиций Диффузия всегда направлена от большей концентрации данного вещества к меньшей, или от системы, обладающей большей свободной энергией к системе с меньшей свободной энергией.Свободной энергией называют ту часть энергии системы, которая может быть использована на какую-то работу. Свободная энергия, отнесенная к молю вещества, носит название химического потенциала. Таким образом Химический потенциал - это мера энергии, которую данное вещество использует на реакции или движение. Вследствие этого передвижение вещества идет от большего химического потенциала к меньшему. Для воды вводится понятие водного потенциала - φ (греч.буква «пси»). Водный потенциал - это способность воды в клетке совершать работу по сравнению с той работой, которую совершала бы чистая вода. φвод.кл Наибольший водный потенциал у чистой, дистиллированной воды. Его принимают за 0. Добавление к воде любых растворимых веществ снижает химический потенциал воды и поэтому водный потенциал любого раствора и биологических жидкостей будет меньше, чем у дистиллированной.воды, т.е. меньше 0, а значит φвод.кл величина отрицательная. Водный потенциал клетки показывает на сколько активность воды в клетке меньше, чем активность чистой воды. Водный потенциал клетки определяется присутствием растворенного вещества - осмотическим потенциалом - φосм и потенциалом, связанным с противодавлением клеточной оболочки - потенциалом давления - φдавл. Осмотический потенциал отражает влияние на водный потенциал частиц растворенных веществ, т.к. частицы растворенных веществ снижают потенциал воды, то φосм - величина отрицательная. Потенциал давления - вода под давлением оболочки обладает большей энергией, чем в отсутствии давления. Молекула воды и растворенного вещества сближаются друг с другом, что приводит к увеличению энергии системы. Следовательно, φд - величина положительная. Таким образом: (-φвод.кл) = (-φосм) + φдавл . Чем меньше клетка насыщена водой, тем более отрицательный водный потенциал. Или: Чем больше концентрация ВЕЩЕСТВ в клетке, тем более отрицательный водный потенциал. Вода всегда передвигается в сторону более отрицательного водного потенциала. Вода движется по градиенту водного потенциала, то есть из области с менее отрицательным водным потенциалом в область с более отрицательным водным потенциалом Проведем параллель между обозначениями водного режима клетки по классической S = Р - Т и термодинамической (-φв.кл) = (-φосм) + φд теории: Водному потенциалу клетки (-φв.кл) соответствует сосущая сила S, различаются только знаком: S - величина положительная и выражается в атмосферах. S = -φ в.кл Осмотический потенциал - осмотическому давлению: Р = -φосм Потенциал давления соответствует тургорному давлению: Т = φд ИЗМЕНЕНИЕ водного потенциала в условиях различного водоснабжения клетки 1. В условиях нормального водоснабжения клетки: (-φв.кл) = (-φосм) + φд . 2. Если клетка находится в состоянии тургора то: φв.кл.→0; φосм=φд . 3. При погружении клетки в гипертонический раствор (осмотический потенциал раствора более отрицательный, чем в клетке), наступает плазмолиз. В этом состоянии: φв.кл.= 0; φв.кл=φосм . 4. При циторризе - φв.кл.= (-φосм) + (-φд) , т.к. клеточная стенка прогибается и φд становится отрицательным. 5. При набухании семян появляется «матричный потенциал» (-φмат) , Он всегда отрицательный; - φв.кл. = (-φосм) + φд + (-φмат) Влияние различных условий на водный потенциал клетки Экологические группы растений: более отрицательный водный потенциал у растений засушливых мест обитания и галофитов (до -200 бар), в то время, как у водных растений = - 1 бар, то есть менее отрицательный. Жизненная форма: боле отрицательный водный потенциал у деревьев, затем кустарники, травы. Чем выше должна подниматься вода, тем более отрицательный водный потенциал должен быть в клетках. Уровень расположения клеток: в клетках листа водный потенциал более отрицательный, чем в клетках корня того же растения. Растение в определенной степени регулирует величину водного и осмотического потенциал: ферментативное превращение сложных нерастворимых веществ в растворимые (крахмала в сахара, белков в аминокислоты) приводит к возрастанию концентрации клеточного сока и повышению величины осмотического потенциала. Задача 1. 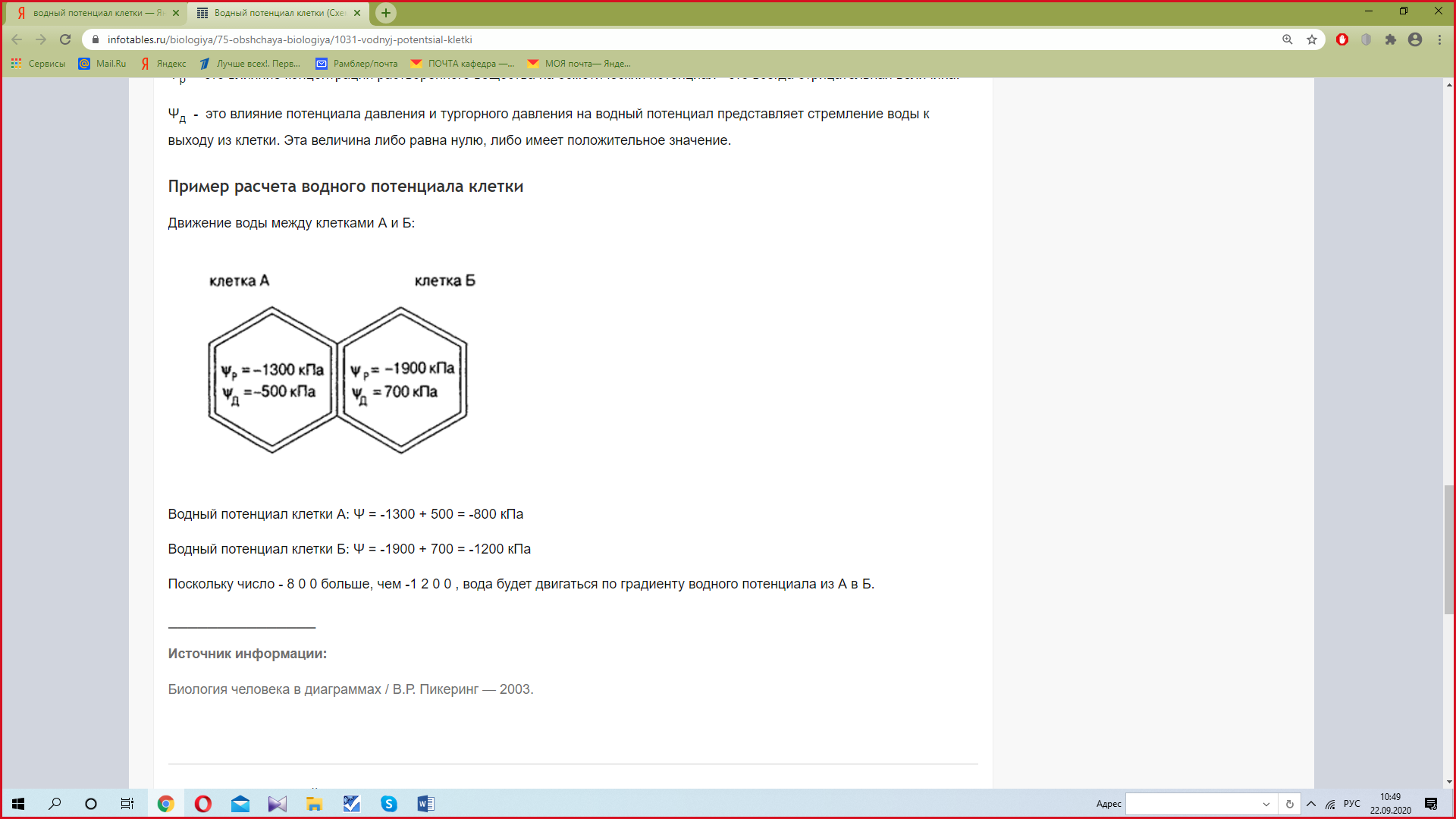 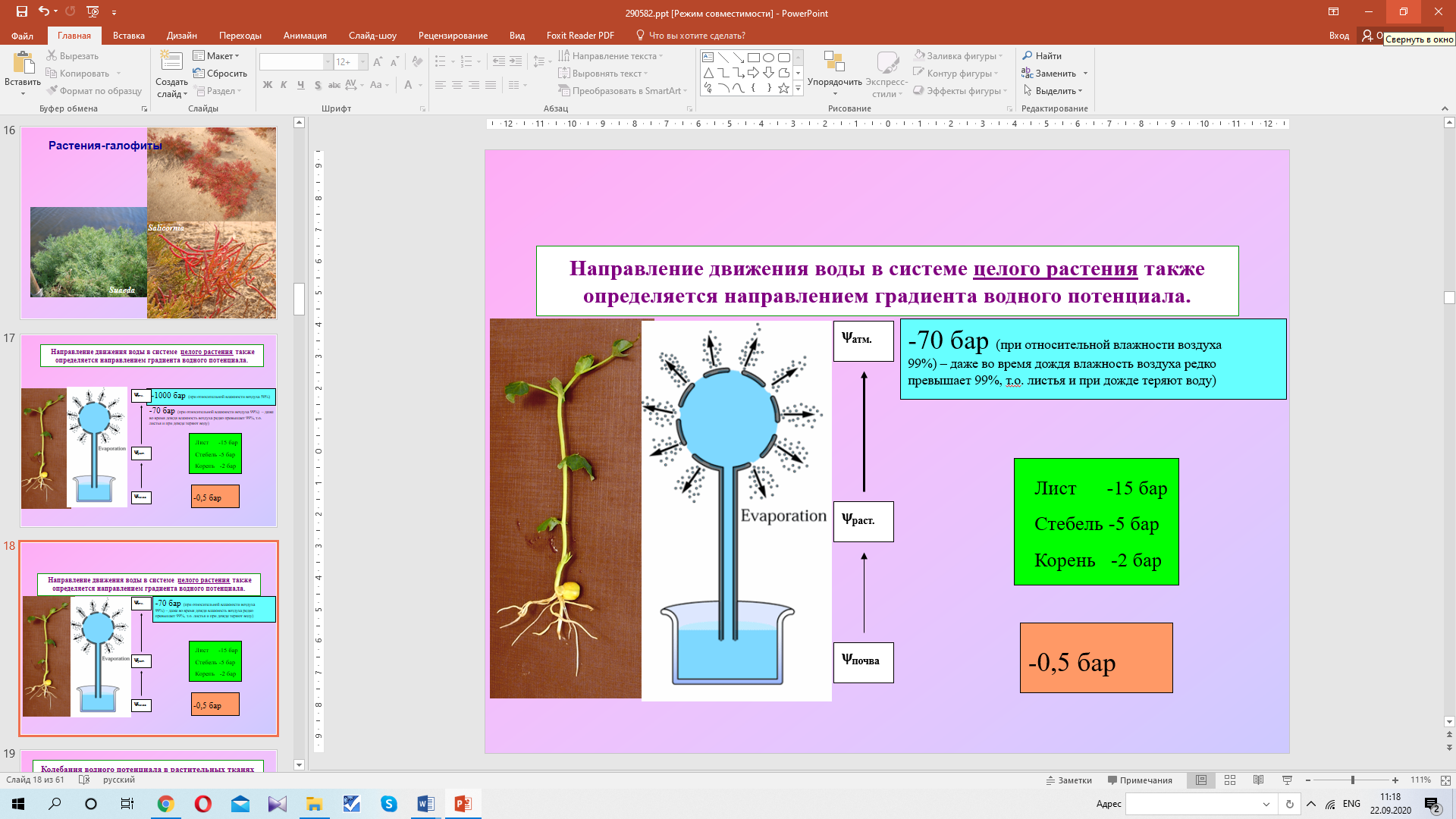 ВИДЕОФРАГМЕНТЫ по теме: Поступление воды в клетку» Как вода проходит через мембрану клетки https://youtu.be/EOkD_E3G3jg Вода и её роль в жизнедеятельности клетки https://youtu.be/iGjfoiYaG4M Осмос. Осмотические явления https://youtu.be/mh6RgoAoXXM Диффузия и осмос. Мембранный транспорт https://youtu.be/z-YzrsAPvL0 Просто о сложном: Что такое осмос и с чем его едят? (опыт с брусочками картофеля) https://youtu.be/HJwNjAj7sUs Определение водного потенциала по изменению размеров клетки (метод Уршпрунга с брусочками картофеля) https://youtu.be/QbNNdb4TH2Q Л3 Физиология растений |