рабочая тетрадь клетка. Материалы для подготовки к занятию Лекция. Введение в физиологию клетки Материалы для подготовки Строение клетки
 Скачать 1.1 Mb. Скачать 1.1 Mb.
|

Модуль №2 Физиология возбудимых клетокЗанятие №3. Общая физиология возбудимых клеток. Морфофункциональная характеристика нервных клеток. Материалы для подготовки к занятию: Лекция. Нейрон Лекция. Общие свойства возбудимых клеток Лекция. Распространение возбуждения Материалы для подготовки возбудимые клетки Материалы для подготовки физиология нервных проводников Учебный фильм - Нервная клетка Учебный фильм - опыты Гальвани Вопросы для самоконтроля:
Домашнее задание:
Физиологические свойства мембран Возбудимость ,проводимость, рефрактерность, лабильность 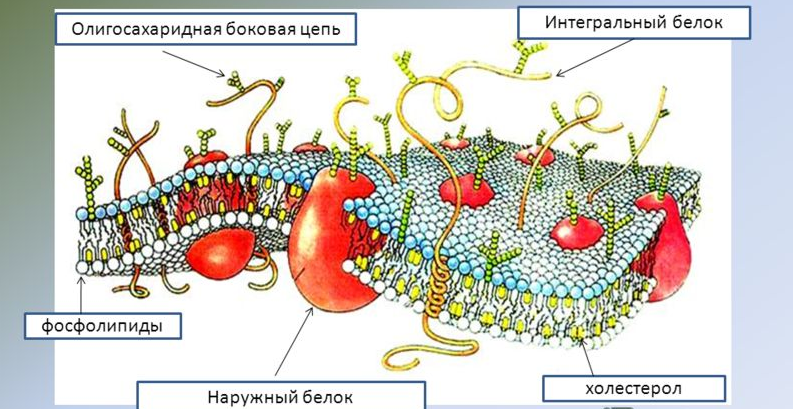
Уравнение Нернста позволяет предсказать максимальный рабочий потенциал, который может быть получен в результате электрохимического взаимодействия, когда известны давление и температура. Таким образом, этот закон связывает термодинамику с электрохимической теорией в области решения проблем, касающихся сильно разбавленных растворов.
Если в формулу Нернста подставить числовые значения констант
Возбудимость – это способность ткани отвечать на раздражение изменением ряда своих свойств. Показатель возбудимости – порог раздражения. Это минимальное по силе раздражение, способное вызвать видимую ответную реакцию ткани. Возбуждение – это в физиологии под возбуждением понимают ответ ткани на раздражение, проявляющийся помимо неспецифических реакций в выполнении специфической для этой ткани функции
К возбудимым тканям относятся: 1. Нервная. 2. Мышечная. 3. Железистая.
Меры возбудимости: 1.Порог раздражения. Это минимальное по силе раздражение, способное вызвать видимую ответную реакцию ткани. 2.Проводимость – способность ткани проводить возбуждение по всей своей длине. Показатель проводимости – скорость проведения возбуждения. Проводимость напрямую зависит от возбудимости ткани: чем выше возбудимость, тем выше проводимость, так как быстрее возбуждается расположенный рядом участок ткани. 3.Рефрактерность – способность ткани терять или снижать возбудимость в процессе возбуждения. При этом в ходе ответной реакции ткань перестает воспринимать раздражитель. Показатель рефрактерности (рефрактерный период) - время, в течение которого возбудимость ткани снижена. Рефрактерный период тем короче, чем выше возбудимость ткани 4.Лабильность – способность ткани генерировать определенное число волн возбуждения в единицу времени в точном соответствии с ритмом наносимого раздражения. Лабильность определяется продолжительностью рефрактерного периода (чем короче рефрактерный период, тем больше лабильность Порог силы – это та минимальная сила раздражителя, которая впервые вызывает видимые ответные реакции, устанавливает зависимость ответной реакции от силы раздражителя. Порог времени Возникновение распространяющегося возбуждения зависит не только от силы раздражителя, но и от времени, в течение которого он действует. Чем больше по силе раздражитель, тем меньшее время он должен действовать для возникновения возбуждения Хронаксия – это минимальное время, требуемое для возбуждения мышечной либо нервной ткани постоянным электрическим током удвоенной пороговой силы (реобаза) Пороговый потенциал – это минимальный сдвиг мембранного потенциала, при котором деполяризация мембраны достигает критического уровня, достаточного для возникновения распространяющегося потенциала действия.
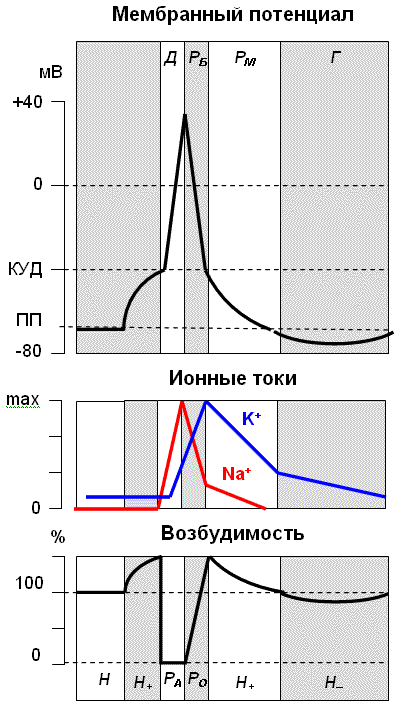 Изменение мембран-ного потенциала, интенсивности калиевого и натриевого трансмембранного тока и возбудимости клетки в разные фазы потенциала действия. Д – фаза деполяризации, Рб – фаза быстрой реполяризации, Рм – фаза медленной реполяризации, Г – фаза гиперполяризации; Н – период нормальной возбудимости, Ра – период абсолютной рефрактерности, Ро – период относительной рефрактерности, Н+ – период супернормальной возбудимости, Н- – период субнормальной возбудимости
Лабильность - это способность ткани генерировать определенное число волн возбуждения в единицу времени в точном соответствии с ритмом наносимого раздражения. Лабильность определяется продолжительностью рефрактерного периода (чем короче рефрактерный период, тем больше лабильность. Измеряется количеством импульсов в секунду.
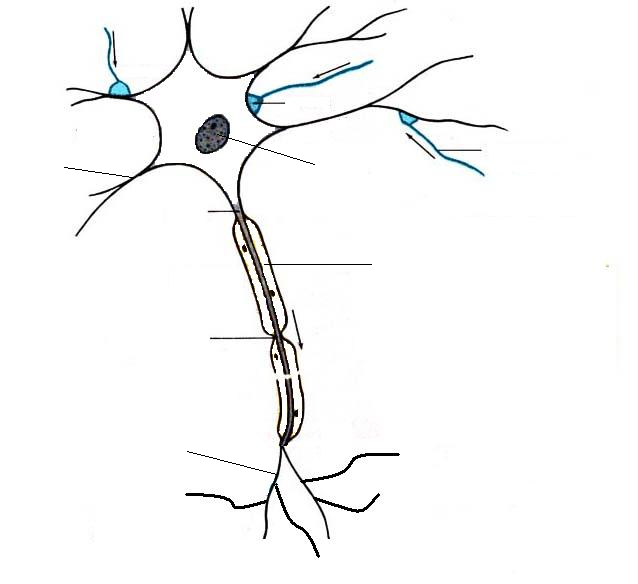 синапс Дендрит Ядро Аксон Миелиновая оболочка Перехват Ранвье Нервные окончания аксона Физиологические свойства нейрона: 1.Раздражимость 2. Раздражение 3. Возбудимость 4. Возбуждение
В зависимости от количества отростков нейроны делят на: 1. Псевдоуниполярные 2. Униполярные 3. Биполярные 4. Мультиполярные В зависимости от функции нейроны делят на: 1. Афферентные 2. Ассоциативные 3. Эфферентые
1. Анатомическая и физиологическая непрерывность волокна 2. Двустороннее проведение возбуждения 3. Изолированное проведение возбуждения в нервных стволах 4. Бездекрементное проведение Практические работы Работа №1 Мембранный потенциал покоя Виртуальный практикум «LupraFisim» Потенциал мышечной клетки, которая в последнее время не подвергалась стимуляции, называется мембранным потенциалом покоя. Он представляет такое состояние, при котором положительные заряды преимущественно распределены на наружной поверхности мембраны, а отрицательные заряды – на ее внутренней поверхности. Цель: продемонстрировать и измерить потенциал покоя на уровне мышечного волокна. Принцип действия: электроды вольтметра подключают к внутренней среде мышечного волокна и к его поверхности. Ход работы. 1. Щелкните кнопку «ВВЕСТИ ЭЛЕКТРОДЫ». 2. Наблюдайте за экраном вольтметром и обратите внимания на изменения мембранного потенциала. 3. Определите величину разности потенциала на экране вольтметра. 4. Для того, чтобы еще раз провести этот эксперимент, щелкните клавишу «ИЗВЛЕЧЬ ЭЛЕКТРОДЫ» РЕЗУЛЬТАТЫ: Разность составила 75 мВ ВЫВОД: Мембранный потенциал покоя увеличился Работа №2 Потенциал действия. Виртуальный практикум «LupraFisim» Когда импульс попадает в мышечное волокно, мембранный потенциал покоя сменяется мембранным потенциалом действия, при этом мембрана мышечного волокна становится гиперпроницаемой для ионов натрия. Ионы натрия в большом количестве поступают в мышечное волокно, при этом распространение электрических зарядов, характерное для потенциала покоя, нарушается (внутри мышечного волокна теперь можно обнаружить больше положительных зарядов). Цель: продемонстрировать и измерить мембранный потенциал действия на уровне мышечного волокна. Принцип действия: два электрода подсоединяют к поверхности мышцы и посылают электрический стимул. Ход работы. 1. Нажмите кнопку «СТИМУЛ». 2. Обратите внимание на то, как формируется деполяризованная волна, и как она движется. 3. Понаблюдайте за экраном вольтметра и обратите внимание на изменения потенциала мембраны. 4. Определите величину потенциала действия. 5. Для повторения эксперимента нажмите кнопку «ПЕРЕЗАПУСК ЭКСПЕРИМЕНТА». РЕЗУЛЬТАТЫ: За один импульс через мембрану проходит небольшая часть ионов Na+ и К+, и концентрационные градиенты обоих ионов сохраняются (в клетке выше уровень К+, а вне клетки выше уровень Na+). Поэтому по мере получения клеткой новых импульсов процесс локального обращения знака мембранного потенциала может повторяться многократно. Распространение потенциала действия по поверхности нервной клетки основано на том, что локальное обращение мембранного потенциала стимулирует открывание соседних потенциал-управляемых ионных каналов, в результате чего возбуждение распространяется в виде деполяризационной волны на всю клетку. Потенциал покоя -60 мВ скачком изменяется на +30 мВ и спустя 1 мс принимает исходное значение ВЫВОД: Ответьте на вопросы: 1. Перечислите фазы ПД, опишите движение ионов в каждую из фаз А) Деполяризация При действии деполяризующего раздражителя на клетку (медиатор, электрический ток) начальная частичная деполяризация клеточной мембраны происходит без изменения ее проницаемости для ионов. Поэтому, несмотря на наличие движущей силы (концентрационный и электрический градиенты), движение Na+ в клетку через быстрые потенциал чувствительные Na-каналы отсутствует. Клетка внутри заряжена отрицательно (разноименные заряды притягиваются друг к другу), а концентрация Na+ вне клетки в 10—12 раз больше, чем внутри клетки. Условием же, обеспечивающим вход Na+ в клетку, является увеличение проницаемости клеточной мембраны, которая определяется состоянием воротного механизма Na-каналов Длительность пребывания электроуправляемого канала в открытом состоянии зависит от величины мембранного потенциала. Суммарный ток ионов в любой момент определяется числом открытых каналов клеточной мембраны и наличием электрохимических градиентов ионов. Когда деполяризация клетки достигает критической величины (Екр., критический уровень деполяризации — КУД), которая обычно составляет 50 мВ (возможны и другие величины), проницаемость мембраны для Na+ резко возрастает — открывается большое число потенциалзависимых Na-каналов и Na+ лавиной устремляется в клетку. Через один открытый Na-канал за 1 мс проходит до 6000 ионов. В результате интенсивного тока Na+ внутрь клетки процесс деполяризации проходит очень быстро. Развивающаяся де-поляризация клеточной мембраны вызывает дополнительное увеличение ее проницаемости и, естественно, проводимости Na+ — открываются все новые и новые активационные Na-каналов. Б) Быстрая реполяризация. Рост ПД прекращается вследствие закрытия натриевых инактивационных ворот и прекращения поступления Na+ в клетку и открытия ворот К-каналов, ведущего к резкому возрастанию выхода К+ из клетки . Препятствуют также росту пика ПД электрический градиент (клетка внутри в этот момент заряжена положительно), а также выход К+ из клетки по каналам утечки, вследствие чего уменьшается число положительно заряженных ионов в клетке. Заряд клетки снова начинает уменьшаться. К+ выталкивается положительным зарядом из клетки и притягивается отрицательным зарядом снаружи клетки. Так продолжается до полного исчезновения положительного заряда внутри клетки. Калий выходит из клетки не только по управляемым каналам, ворота которых открыты, но и по неуправляемым — каналам утечки, что несколько замедляет ход восходящей части ПД и ускоряет ход нисходящей составляющей ПД. В) Медленная реполяризация. Связана с тем, что проницаемость клеточной мембраны для К+ все еще высока (активационные ворота калиевых каналов открыты), К+ продолжает быстро выходить из клетки согласно концентрационному градиенту. Поскольку клетка теперь уже снова внутри имеет отрицательный заряд, а снаружи — по-ложительный, электрический градиент препятствует выходу К+ из клетки, что снижает его проводимость, хотя он продолжает выходить. Это объясняется тем, что действие концентрационного градиента выражено значительно сильнее электрического градиента. Таким образом, вся нисходящая часть пика ПД обусловлена выходом К+ из клетки. Замедление реполяризации объясняется уменьшением проницаемости клеточной мембраны для К+ и замедлением выхода его из клетки из-за закрытия значительной части ворот К-каналов. Вторая причина замедления тока К+ из клетки связана с возрастанием положительного заряда наружной поверхности клетки и формированием противоположно направленного электрического градиента. Таким образом, главную роль в возникновении ПД играет Na+, входящий в клетку при повышении проницаемости клеточной мембраны и обеспечивающий всю восходящую часть пика ПД. Однако проницаемость мембраны для К+ тоже играет важную роль. Г) Гиперполяризация. Обычно является результатом еще сохраняющейся повышенной проницаемости клеточной мембраны для К+. Активационные ворота К-каналов еще не полностью закрыты, поэтому К+ продолжает выходить из клетки согласно концентрационному градиенту, что и ведет к гиперполяризации клеточной мембраны. Постепенно проницаемость клеточной мембраны возвращается к исходной (натриевые и калиевые ворота возвращаются в исходное состояние), а мембранный потенциал становится таким же, каким он был до возбуждения клетки. Na/K-помпа непосредственно за фазы потенциала действия не отвечает, хотя она работает непрерывно в покое и продолжает работать во время развития ПД. Потенциал действия развивается благодаря движению ионов с огромной скоростью согласно концентрационному и частично электрическому градиентам. Возможно, Na/K-помпа способствует развитию следовой гиперполяризации. В некоторых клетках, например в тонких немиелинизированных нервных волокнах (болевых афферентах), хорошо выражена длительная следовая гиперполяризация. Она обеспечивается работой Na/K-насоса, активируемого процессом возбуждения (накопившимся в клетке Na+: на каждые 2К+, возвращаемых в клетку, выводится 3Na+ из клетки). Если блокировать выработку энергии, то эта гиперполяризация исчезает. 2.Опишите изменение заряда на мембране в процессе генерации ПД. Вход Na+не только нейтрализует отрицательный заряд, имеющийся исходно на внутренней стороне мембраны, до нуля, но и перезаряжает ее до положительных значений порядка +20 — +40 мВ. Затем Na-проницаемость снижается до исходных низких значений. Во вторую фазу потенциала действия резко возрастает проницаемость мембраны к ионам калия, концентрация которых выше в цитоплазме клетки, чем в наружной среде, и они выходят из клетки по концентрационному градиенту, вынося положительные заряды и способствуя их накоплению на наружной стороне мембраны. Это сопровождается одновременным накоплением анионов на внутренней поверхности мембраны и реполяризацией мембраны вплоть до исходных значений заряда и потенциала (возвращение к состоянию покоя). Важнейшими условиями для открывания натриевых каналов и генерации потенциал действия является деполяризация клетки до определенного уровня, называемого критическим, или «пороговым». Только по достижении «пороговой» величины мембранного потенциала (порядка -50 — -30 мВ) происходит открывание натриевых, а затем и калиевых каналов, и начинается генерация потенциала действия. Другой важнейшей особенностью генерации потенциала действия является существование короткого периода абсолютной и относительной невозбудимости мембраны: в короткий период порядка 1-2 мс после прекращения генерации потенциала действия не удается вызвать новую генерацию потенциала действия. Этот период кратковременной нечувствительности мембраны, называемый рефрактерным периодом, определяется инактивированным состоянием натриевых каналов во вторую, реполяризационную фазу генерации потенциала действия. Ионные каналы вновь приобретают способность открываться в ответ на пороговую деполяризацию лишь после возвращения мембранного потенциала к уровню потенциала покоя. Генерация потенциала действия является не внешним проявлением, а самой сутью феномена возбуждения клетки. Именно с помощью потенциала действия нейроны получают, перерабатывают и передают биологически важную информацию из внешней среды, а также от одной клетки к другой, а мышечные клетки начинают сокращаться, а значит, обеспечивается двигательная активность органов, стенки которых состоят из возбудимых гладкомышечных клеток: сердца, сосудов, пищеварительного тракта. В железистых клетках потенциал действия запускает процесс секреции. 3.Объясните взаимосвязь изменения заряда мембраны и возбудимости. При действии на возбудимую ткань раздражителя пороговой или надпороговой силы происходит кратковременное изменение заряда на участке клеточной мембраны, которое называется ток действия или потенциал действия. Потенциал действия связан с изменением ионной проницаемости мембраны. Если сила раздражителя не достигает пороговых значений, открывается часть натриевых каналов, натрий входит внутрь клетки. Это приводит к снижению деполяризации мембраны, но соотношение зарядов между наружной и внутренней поверхностью клеточной мембраны быстро восстанавливается. Такое изменение потенциала называется локальный ответ. При достижении пороговой силы стимула все Na+ каналы открываются, и Na+ лавиной устремляется внутрь клетки, заряд на мембране уменьшается до 0. Эта фаза называется деполяризация . Из-за поступления большого количества положительных ионов во внутриклеточное пространство количество положительных зарядов на внутренней поверхности мембраны становится больше, чем снаружи, то есть происходит смена мембранного заряда на противоположный (реверсия). Работа №3 Демонстрация воздействия анестезирующих средств и низкой температуры на генерацию и проведение ПД Виртуальный практикум «LupraFisim» Цель: Оценить влияние некоторых анестетиков и воздействие низкой температуры на возбудимость и скорость проводимости нерва. Описание эксперимента: седалищный нерв лягушки некой точке подвергается воздействию стимулирующего импульса при следующих условиях: - после того, как нерв был смочен лидокаином (блокирующим каналы натрия); - после того, как нерв был смочен эфиром; - после того, как на нерв было помещено несколько льдинок. Ход работы.
РЕЗУЛЬТАТЫ: При воздействии электрическим стимулом на нерв ответная реакция была положительной время измерения 0,1 мс; скорость проводимости 400 м/с. Скорость проводимости при= 0,06мс. При воздействии электрическим стимулом на нерв, смоченный в растворе лидокаина и эфира ответной реакции не было. При воздействии электрическим стимулом на нерв с помещенными на него льдинками ответная реакция была положительной, время измерения 0,1 мс, скорость проводимости D2/T2. Скорость проводимости = 8,8мс ВЫВОД: Отсутствие ответной реакции обусловлено угнетением нервной проводимости за счет блокады натриевых каналов в нервных окончаниях и нервных волокнах, низкая температура значительно снижает ответную реакцию. Работа №4 Определение порога силы и демонстрация явления суммации Виртуальный практикум «LupraFisim» Порог силы – это минимальная интенсивность единичного стимула, при которой возникает и распространяется потенциал действия. Применение нескольких подпороговых стимулов с высокой частотой приводит к появлению потенциала действия в результате суммации. Принцип действия: Нерв подвергается воздействию электрических раздражителей все большей силы, пока не возникнет потенциал действия. Затем нерв подвергается воздействию нескольких подпорговых раздражителей с высокой частотой. Ход работы:
РЕЗУЛЬТАТЫ: Возникновение потенциала действия происходит при интенсивности стимула 3 мВ и числе стимулов равном 1. Аналогично при интенсивности стимула 2,8 мВ и числе стимулов – 4. При снижении порога возбудимости до 2,3 мВ и увеличении числа стимулов до 600 ПД не был получен. ВЫВОД: 1)укажите зависимость уровня возбудимости от величины порога силы; 2) перечислите временные пороги возбудимости; 3) поровый потенциал, как электрофизиологический критерий (порог) возбудимости Чем ниже порог силы тем выше возбудимость ткани. Раздражители, сила которых ниже порога возбуждения, рассматриваются как подпороговые. Если сила раздражения превосходит порог возбуждения, величина ответной реакции ткани (возбуждения) возрастает вплоть до известного, определенного для каждого живого образования предела. Дальнейшее увеличение силы раздражителя уже не ведет к росту ответной реакции. Минимальная сила раздражителя, вызывающая наибольший (максимальный) ответ ткани, называется максимальной силой раздражения. Раздражители, сила которых меньше или больше максимальной, называются, соответственно, субмаксимальными и супермаксимальными. Пороговый потенциал – это минимальная величина, на которую надо уменьшить потенциал покоя, чтобы вызвать возбуждение (потенциа действия). В данном случае пороговым является потенциал равный 3,0 мВ. Для того чтобы вызвать ответную реакцию меньшим (подпороговым) потенциалом, необходимо увеличить частоту импульсов. Работа №5 Определение скорости проводимости Виртуальный практикум «LupraFisim» Цель: измерить скорость проводимости нерва, с использованием следующих типов нервов: - тонкий миелинизированный нерв лягушки; - немиелинизированный нерв крысы; - толстый миелинизированный нерв крысы. Принцип действия: воздействию электрического раздражителя подвергаются нервы разного типа, и определяется скорость их проводимости: с помощью двух электродов, размещенных на известном расстоянии от электрода-раздражителя, замеряется потенциал действия. Так как расстояние известно, то, засекая время, можно вычислить скорость проводимости. Ход работы.
РЕЗУЛЬТАТЫ: Скорость проводимости седалищного нерва лягушки: Значение интенсивности стимула – 3 мВ; Время – 6,133 мс Скорость – 6,52 м/с. Скорость проводимости безмиелинового нерва крысы: Значение интенсивности стимула – 2,5 мВ; Время – 6,36 мс Скорость – 6,28 м/с. Скорость проводимости миелинового нерва крысы: Значение интенсивности стимула – 3,0 мВ; Время – 5, 6 мс Скорость – 7,14 м/с. ВЫВОД: наличие миелинового слоя напрямую влияет на скорость проводимости нерва, чем он толще, тем выше скорость проводимости Занятие №4. Морфо-функциональная характеристика мышечных клеток. Классификация мышечных клеток, роль в организме. Механизм мышечного сокращения. Материалы для подготовки к занятию: Материалы для подготовки физиология мышц Презентация Физиология мышц Вопросы для самоконтроля: 1. Виды мышц в организме, Физиологические свойства мышечной ткани. 2. Морфо-функциональная характеристика скелетных мышц. 3. Механизм мышечного сокращения. Роль кальция в мышечном сокращении. 4. Виды и режимы мышечных сокращений. 5. Одиночное мышечное сокращение скелетной мышцы, условия получения, фазы. Временные соотношения возбуждения и сокращения в мышцах разных видов. 6. Тетаническое сокращение. Условия получения различных видов тетануса. 7. Особенности строения и физиологических свойств гладкой мышцы. Автоматия, определение понятия, значение. Домашнее задание:
Классификация мышц по строению: 1. Поперечнополосатые (скелетные) 2. Гладкие Поперечнополосатые мышцы делят на: 1. Агонисты 2. Антагонисты Перечислите функции скелетных мышц: Обеспечивает возможность выполнения произвольных движений. Сокращающаяся мышца воздействует на кости или кожу, к которым она прикрепляется. При этом один из пунктов прикрепления остаётся неподвижным — так называемая точка фиксации (лат. púnctum fíxsum), которая в большинстве случаев рассматривается в качестве начального участка мышцы. Перемещающийся фрагмент мышцы называют подвижной точкой, (лат. púnctum móbile), которая является местом её прикрепления. Тем не менее, в зависимости от выполняемой функции, punctum fixum может выступать в качестве punctum mobile, и наоборот.
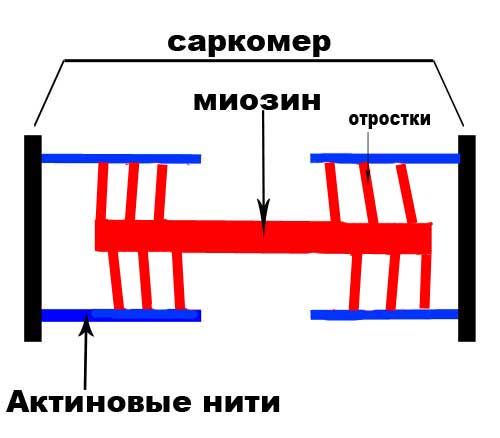 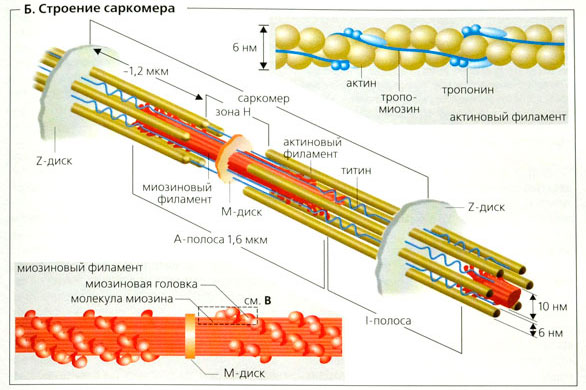 Этапы мышечного сокращения: Выделяют несколько последовательных этапов запуска и осуществления мышечного сокращения. 1. Потенциал действия распространяется вдоль двигательного нервного волокна до его окончаний на мышечных волокнах. 2. Каждое нервное окончание секретирует небольшое количество нейромедиатора ацетилхолина. 3. Ацетилхолин действует на ограниченную область мембраны мышечного волокна, открывая многочисленные управляемые ацетилхолином каналы, проходящие сквозь белковые молекулы, встроенные в мембрану. 4. Открытие управляемых ацетилхолином каналов позволяет большому количеству ионов натрия диффундировать внутрь мышечного волокна, что ведет к возникновению на мембране потенциала действия. 5. Потенциал действия проводится вдоль мембраны мышечного волокна так же, как и по мембране нервного волокна. 6. Потенциал действия деполяризует мышечную мембрану, и большая часть возникающего при этом электричества течет через центр мышечного волокна. Это ведет к выделению из саркоплазматического ретикулума большого количества ионов кальция, которые в нем хранятся. 7. Ионы кальция инициируют силы сцепления между актиновыми и миозиновыми нитями, вызывающие скольжение их относительно друг друга, что и составляет основу процесса сокращения мыщц. 8. Спустя долю секунды с помощью кальциевого насоса в мембране саркоплазматического ретикулума ионы кальция закачиваются обратно и сохраняются в ретикулуме до прихода нового потенциала действия. Удаление ионов кальция от миофибрилл ведет к прекращению мышечного сокращения. Роль кальция в сокращении: В расслабленном состоянии концы актиновых нитей, отходящие от двух последовательных Z-дисков, лишь незначительно перекрываются. Наоборот, в сокращенном состоянии актиновые нити втягиваются внутрь между миозиновыми так сильно, что их концы максимально перекрывают друг друга. При этом Z-диски притягиваются актиновыми нитями к концам миозиновых. Таким образом, мышечное сокращение осуществляется путем механизма скольжения нитей. Что заставляет нити актина скользить внутрь среди нитей миозина? Это связано с действием сил, генерируемых при взаимодействии поперечных мостиков, исходящих от нитей миозина, с нитями актина. В условиях покоя эти силы не проявляются, однако распространение потенциала действия вдоль мышечного волокна приводит к выделению из саркоплазматическо-го ретикулума большого количества ионов кальция, которые быстро окружают миофибриллы. В свою очередь, ионы кальция активируют силы взаимодействия между нитями актина и миозина, в результате начинается сокращение. Для осуществления процесса сокращения необходима энергия. Ее источником являются высокоэнергетические связи молекулы АТФ, которая разрушается до АДФ с высвобождением энергии.
График потенциала действия: 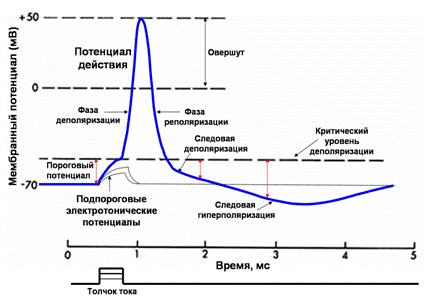 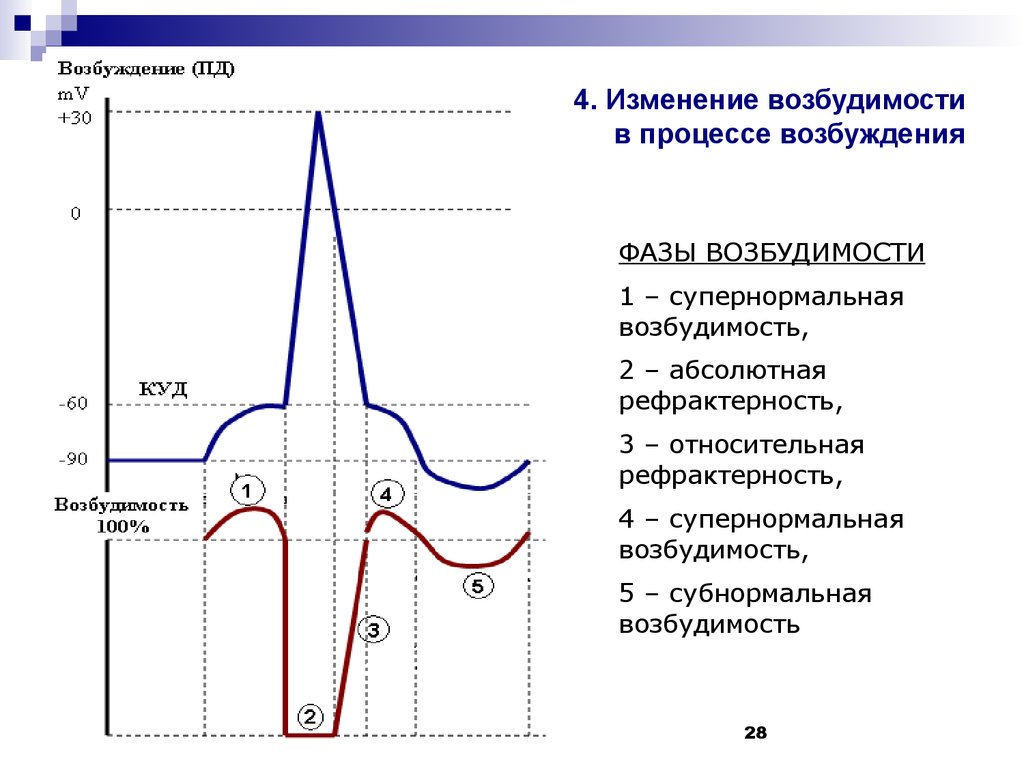 Рис А Головки миозина присоединяются к центрам связывания актинового филламента . Рис. Б Взаимодействие миозина с актином приводит к конформационным перестройкам молекулы миозина. Головки приобретают АТФазную активность и поворачиваются на 120 ° . За счет поворота головок нити актина и миозина передвигаются на «один шаг» друг относительно друга Рис. В Рассоединение актина и миозина и восстановление конформации головки происходит в результате присоединения к головке миозина молекулы АТФ и ее гидролиза в присутствии Са++ Рис. Г Цикл «связывание – изменение конформации – рассоединение – восстановление конформации» происходит много раз, в результате чего актиновые и миозиновые филламенты смещаются друг относительно друга, Z -диски саркомеров сближаются и миофибрилла укорачивается 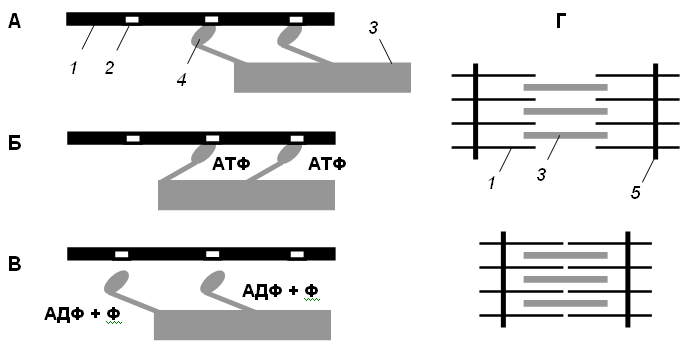 1 – актиновый филламент, 2 – центр связывания, 3 – миозиновый филламент, 4 – головка миозина, 5 – Z -диск саркомера. В состоянии покоя скольжения нитей в миофибрилле не происходит, так как центры связывания на поверхности актина закрыты молекулами белка тропомиозина (рис. 3, А, Б). Возбуждение (деполяризация) миофибриллы и собственно мышечное сокращение связаны с процессом элетромеханического сопряжения, который включает ряд последовательных событий. • В результате срабатывания нейромышечного синапса на постсинаптической мембране возникает ВПСП, который генерирует развитие потенциала действия в области, окружающей постсинаптическую мембрану. • Возбуждение (потенциал действия) распространяется по мембране миофибриллы и за счет системы поперечных трубочек достигает саркоплазматического ретикулума. Деполяризации мембраны саркоплазматического ретикулума приводит к открытию в ней Са++ -каналов, через которые в саркоплазму выходят ионы Са++ (рис. 3, В). • Ионы Са++ связываются с белком тропонином. Тропонин изменяет свою конформацию и смещает молекулы белка тропомиозина, которые закрывали центры связывания актина (рис. 3, Г). • К открывшимся центрам связывания присоединяются головки миозина, и начинается процесс сокращения (рис. 3, Д). Для развития указанных процессов требуется некоторый период времени (10–20 мс). Время от момента возбуждения мышечного волокна (мышцы) до начала ее сокращения называют латентным периодом сокращения . 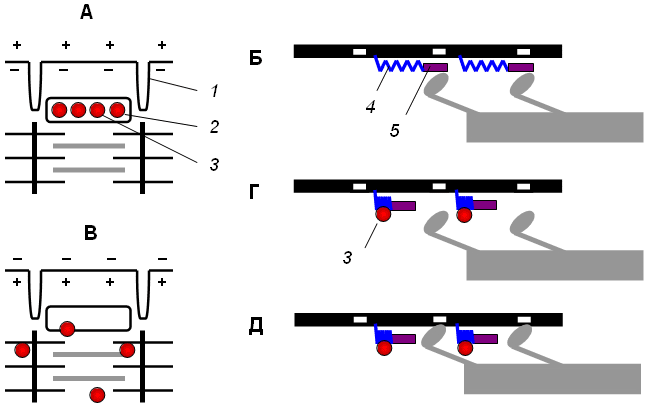 1 – поперечная трубочка саркоплазматичекой мембраны, 2 –саркоплазматичекий ретикулум, 3 – ион Са++ , 4 – молекула тропонина, 5 – молекула тропомиозина.
1. Одиночное 2. Зубчатый тетанус 3. Гладкий тетанус
Режимы: 1.Изотонический 2.Изометрический 3. Ауксотонический
Возникает в том случае, когда каждый последующий импульс приходит в фазу расслабления мышцы.
Каждое последующее раздражение осуществляется в фазу повышенной возбудимости. Тетанус при этом будет максимальным.
Каждое последующее раздражение осуществляется в фазу пониженной возбудимости. Тетанус при этом будет минимальным. Практические работы: Работа №1 Одиночное сокращение скелетных мышц Виртуальный практикум «LupraFisim» Одиночное сокращение скелетных мышц представляет собой ответную реакцию в виде сокращения на одиночный стимул. Цели: 1. Визуализировать и измерить фазы простого сокращения, продемонстрировать связи между силой стимула и силой мышечного сокращения (количеством мышечных волокон, задействованных в ответной реакции). 2. Продемонстрировать влияние низкой температуры на мышечную возбудимость и сократимость. Принцип действия: скелетная мышца подвергается воздействию единичного электрического стимула, при этом получается графическое изображение мышечного сокращения (миограмма) в нормальных условиях искусственного охлаждения мышцы. Ход работы. Практическая работа состоит из двух экспериментальных составляющих:
РЕЗУЛЬТАТЫ: Увеличение силы импульса ведет к постепенному возрастаю амплитуды сокращений мышечного волокна. При достижении силы импульса 8,1 мВ амплитуда мышечного сокращения перестает расти. Длительность латентного периода – 1 мс. Фаза укорочения – 4,9 мс. Фаза расслабления – 15,9 мс. В норме амплитуда мышечного сокращения равна 2,7. При охлаждении параметры меняются следующим образом. Силы импульса 8,1 мВ. Длительность латентного периода – 4,8 мс. Фаза укорочения – 5,2 мс. Фаза расслабления – 13,0 мс. Амплитуда мышечного сокращения равна 1,4. ВЫВОД: Чем сильнее электрический импульс, тем больше амплитуда мышечного сокращения. Причем она возрастает до определенного момента. Охлаждение мышцы значительно снижает амплитуду ее сокращения при тех же значениях импульса, а также увеличивает его длительность. Работа №2 Виды тетанического сокращения. Виртуальный практикум «LupraFisim» Совокупность сокращений представляет собой ответную реакцию в виде сокращения скелетных мышц, которая наступает после применения как минимум двух стимулов до того, как закончится период сокращения, вызванный первым стимулом (15-20 миллисекунд). Цель: изучить виды тетанического сокращения поперечно-полосатых мышц и проанализировать получающиеся миограммы. Принцип действия: примените несколько стимулов разной частоты к скелетным мышцам, регистрируя сокращения на миограммах. Ход работы. Практическая работа состоит из двух частей:
РЕЗУЛЬТАТЫ: Миограмма 1. 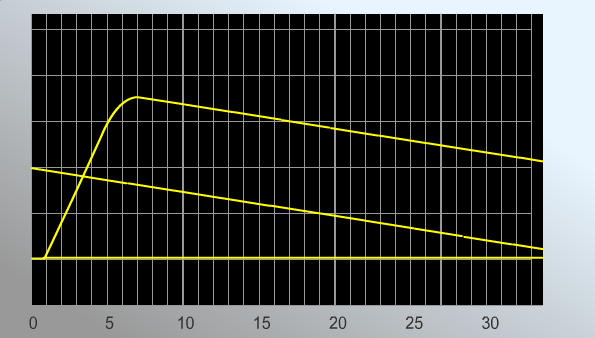 Стимуляция мышцы с частотой 20 стимулов в секунду приводит к возникновению полного тетануса. Верхняя часть миограммы имеет вид гладкого плоскогорья. Миограмма 2. 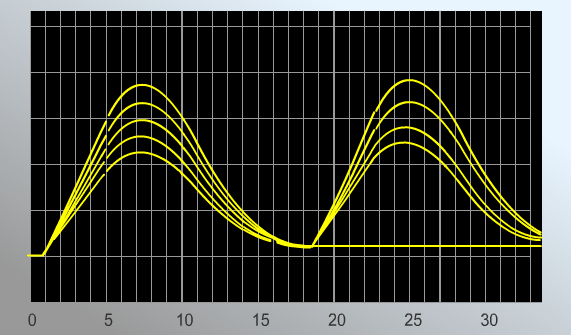 Миограмма 3. 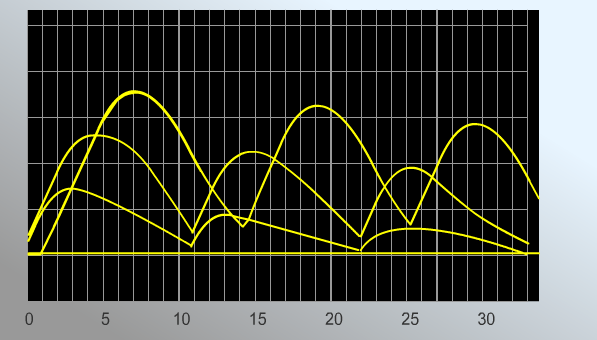 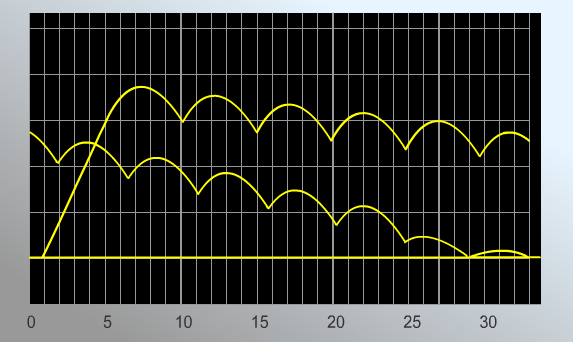 В случае частот 5, 6,5 и 10 стимулов в секунду получается зубчатый тетанус. ВЫВОД:1Перечислите условия получения зубчатого и гладкого тетануса. 2. Назовите условия получения оптимального и пессимального тетануса. В случае воздействия каскада стимулов небольшой частоты (5-10 в с.) получается что каждый второй стимул приходится на фазу расслабления мышечного волокна. График имеет вид зубчатого плоскогорья. При повышении частоты стимуляции каждый второй стимул приходится на фазу сокращения мышечного волокна. График имеет вид гладкого плоскогорья. Амплитуда гладкого тетануса колеблется в широких пределах в зависимости от частоты стимуляции нерва. При некоторой оптимальной (достаточно высокой) частоте стимуляции амплитуда гладкого тетануса становится наибольшей. Такой гладкий тетанус получил название оптимума. При дальнейшем повышении частоты стимуляции нерва развивается блок проведения возбуждения в нервно-мышечных синапсах, приводящий к расслаблению мышцы в ходе стимуляции нерва — пессимум.Частота стимуляции нерва, при которой наблюдается пессимум, получила название пессимальной. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||