Механизмы генетической изменчивости. Полиморфизм белков. Наследственные болезни
 Скачать 77.93 Kb. Скачать 77.93 Kb.
|

|
Федеральное государственное бюджетное образовательное учреждение высшего образования «Новосибирский государственный медицинский университет» Министерства здравоохранения Российской Федерации (ФГБОУ ВО НГМУ Минздрава России) Кафедра медицинской химии Дисциплина «Биохимия» Реферат на тему: «Механизмы генетической изменчивости. Полиморфизм белков. Наследственные болезни» Выполнила: студентка 2 курса 2 группы Педиатрического факультета Специальность:медико-профилактическое дело Мамаева Валерия Евгеньевна Проверил: К.м.н Долганова Ольга Михайловна Подпись___________ Новосибирск , 2022 Содержание Введение.....................................................................................................3 Глава 1.Механизмы генетической изменчивости…...…....…..........4-5 1.1.Мутагенез………………………………………………….............................… 4 1.2.Организация генома человека……….……............................. 5 Глава 2. Полиморфизм белков………………………………….....................….6-8 Глава 3. Наследственные болезни………………...…………..................…..…9 Заключение………………………………………………………...............................….…...11 Список литературы…………………………………………..........................………..….12 Введение Точная работа всех матричных биосинтезов — репликации, транскрипции и трансляции — обеспечивает копирование генома и воспроизведение фенотипических характеристик организма в поколениях, т. е. наследственности. Однако биологическая эволюция и естественный отбор возможны только при наличии генетической изменчивости. Установлено, что геном постоянно претерпевает разнообразные изменения. Несмотря на эффективность механизмов коррекции и репарации ДНК, часть повреждений или ошибок в ДНК остаётся. Изменения в последовательности пуриновых или пиримидиновых оснований в гене, не исправленные ферментами репарации, получили название «мутации». Глава 1. Механизмы генетической изменчивости Существенный вклад в генетическую изменчивость вносят перестройки хромосом в процессе мейоза. Как уже указывалось ранее, слияние яйцеклетки со сперматозоидом у эукариотов сопровождается генетическими рекомбинациями, в ходе которых происходит обмен участками ДНК между гомологичными хромосомами. Это приводит к появлению потомства с новой комбинацией генов. Ген или части генов могут перемещаться из одного места хромосомы в другие. Эти подвижные элементы или фрагменты ДНК получили название транспозонов и ретротранспозонов. Транспозоны — участки ДНК, удаляемые из одного локуса хромосомы и встраиваемые в другой локус той же или другой хромосомы. Ретротранспозоны не покидают исходного положения в молекуле ДНК, но могут копироваться, и копии встраиваются, подобно транспозонам, в новый участок. Включаясь в гены или участки около генов, они могут вызывать мутации и изменять их экспрессию. Геном эукариотов подвергается изменениям и при заражении ДНК- или РНК-содержащими вирусами, которые внедряют свой генетический материал в ДНК клеток хозяина. Мутагенез. Изменения в геноме могут быть разнообразны и затрагивать различные по протяжённости участки ДНК от хромосом и генов до отдельных нуклеотидов. 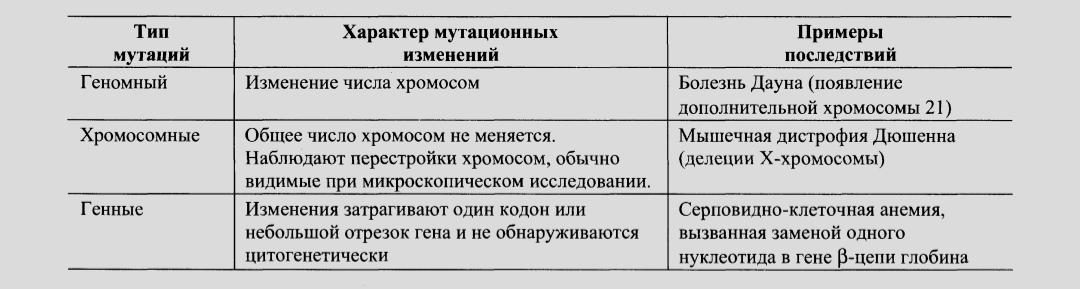 Генные, или точечные, мутации бывают в основном 3 видов: замены, при которых одно азотистое основание в ДНК замещается на другое; вставки, обеспечивающие внедрение в молекулу ДНК одного или нескольких дополнительных нуклеотидов; делении (или выпадения) одного или нескольких нуклеотидов, при которых происходит укорочение молекулы ДНК. Организация генома человека. Геном эукариотических клеток (клеток человека в частности) содержит значительно больше ДНК, чем геном прокариотов.Общая длина ДНК гаплоидного набора из 23 хромосом человека составляет 3,5 x 109 пар нуклеотидов, что в 1000 раз превосходит размер генома прокариотов. Этого количества ДНК достаточно для создания нескольких миллионов генов. Последовательность нуклеотидов в ДНК человека имеет области, кодирующие белки (не более 2% общего генома), области, кодирующие РНК (около 20% генома), и повторяющиеся последовательности (более 50% общего генома). Функции «избыточной» ДНК до конца не ясны, полагают, что она участвует в регуляции экспрессии генов, процессинга РНК, выполняет структурные функции, повышает точность гомологичного спаривания и рекомбинации хромосом в процессе мейоза, способствует успешной репликации. Большая часть этой ДНК возникла в результате обратной транскрипции РНК и благодаря наличию подвижных элементов. Часто повторяющиеся последовательности ДНК Во фракции «избыточной» ДНК выделяют относительно короткие последовательности длиной от 2 до 10 пар нуклеотидов, повторяющиеся миллионы раз. Эти часто повторяющиеся последовательности получили название «сателлитной ДНК». Они составляют около 10% всего генома человека, рассеяны по всему генетическому материалу клетки, но преимущественно локализуются в центромерных и теломерных областях большинства хромосом. Умеренно повторяющиеся последовательности ДНК Выделяют также группу умеренно повторяющихся последовательностей ДНК, очень гетерогенную по длине и числу копий, составляющую более 30% генома человека. Эта фракция включает ДНК, кодирующую структуру рРНК, тРНК и некоторых мРНК. Гены гистонов присутствуют в геноме в количестве нескольких сотен копий и также принадлежат к этому классу. Умеренно повторяющиеся последовательности ДНК включают участки, которые не транскрибируются, но важны в регуляции экспрессии генов: промоторы и энхансеры. В этой группе были обнаружены последовательности ДНК длиной примерно в 300 пар нуклеотидов, которые повторяются около миллиона раз (в среднем через каждые 5000 пар нуклеотидов) и рассеяны по всему геному человека. Уникальные последовательности ДНК Они представлены ДНК-последовательностями, которые присутствуют в геноме в количестве одной или нескольких копий, и транскрибируются, образуя мРНК, содержащие информацию о различных белках. Нередко уникальные последовательности образуют мультигенные семейства, располагающиеся в виде кластеров в определённых областях одной или нескольких хромосом. Примерами мультигенных семейств могут служить гены рибосомальных, транспортных и малых ядерных РНК, гены α- и β-глобинов, тубулинов, миоглобина, актина, трансферина и многих других. Глава 2.Полиморфизм белков Поскольку большинство нормальных клеток человека диплоидны, то они содержат две копии каждой хромосомы, одна из которых получена от отца, а вторая от матери. Эти две копии одной и той же хромосомы называют гомологичными (рис. 4-59). В ДНК каждой хромосомы содержится более тысячи генов. Соответствующие друг другу гены в гомологичных хромосомах называют аллелями. Аллели могут быть идентичными и содержат одинаковую последовательность нуклеотидов. В этом случае индивидуум, имеющий такие аллели, будет гомозиготен по данному признаку. Если аллели различаются по последовательности нуклеотидов в ДНК, то говорят о гетерозиготном наследовании гена. В этом случае индивидуум будет иметь 2 белковых продукта гена, различающихся по аминокислотной последовательности. У каждого человека существует только 2 разных аллеля одного гена, тогда как в популяции людей вариантов аллелей может быть огромное множество. Как уже говорилось ранее, изменчивость структуры ДНК, а, следовательно, разнообразие аллелей, обусловлено мутационным процессом и рекомбинациями в гомологичных хромосомах половых клеток. Если в ходе мейоза рекомбинации сопровождаются обменом участками ДНК, меньшими по размеру, чем ген, то такой процесс может приводить к появлению новых, прежде не существовавших аллелей. А поскольку рекомбинации — более частые события, чем мутации в кодирующих участках гена, то разнообразие вариантов аллелей обусловлено главным образом ими. Существование в популяции 2 и большего числа аллелей одного гена называют «аллеломорфизм», или «полиморфизм», а белковые продукты, образующиеся в ходе экспрессии этих вариантов гена — «полиморфы». Разные аллели встречаются в популяции с разной частотой. К полиморфам относят только те варианты, распространённость которых в популяции не меньше 1%. В процессе эволюции отдельные гены амплифицируют с образованием копий, а их структура и положение могут изменяться в результате мутаций и перемещений не только внутри хромосомы, но и между хромосомами. Со временем это приводит к появлению новых генов, кодирующих белки, родственные исходному, но отличающиеся от него определёнными свойствами и занимающие в хромосомах разные генные локусы (или места). К родственным белкам относят изобелки, представляющие собой варианты белков, выполняющие одну и ту же функцию и обнаруживаемые в пределах одного вида организмов. Так, в группе из 2000 генов человека, кодирующих факторы транскрипции и транскрипционные активаторы, идентифицировано 900, относящихся к семейству белков, имеющих «цинковые пальцы». Существует 46 генов фермента глицеральдегид- 3-фосфатдегидрогеназы, осуществляющего единственную окислительную реакцию в метаболическом пути катаболизма глюкозы до пирувата. Выявлены семейства родственных белков, возникшие в ходе эволюции из одного «предкового» гена, или гена-предшественника. 1. Гемоглобины человека В ходе эволюции из единичных генов-предшественников возникли семейства генов α- и β-глобинов (рис. 4-60), на хромосомах 16 и 11 соответственно. В процессе онтогенеза у людей образуются разные виды гемоглобинов, обеспечивающие наилучшую адаптацию к меняющимся условиям существования. НbЕ — эмбриональный, синтезируется у зародыша в первые месяцы развития, НbЕ — фетальный, обеспечивает дальнейшее внутриутробное развитие плода, а НbА и НbА2 осуществляют транспорт кислорода в организме взрослого человека. Эти белки представляют собой тетрамеры, состоящие из полипептидных цепей двух видов: α и β в НbА (2α2β), α и ε в НbЕ (2α2ε), а у остальных гемоглобинов β-цепи заменены на y-полипептиды в НbF (2α2y) или на 5-цепи в НbА2 (2α2δ). Полиморфизм гемоглобинов в популяции людей очень велик. Наряду с генами, кодирующими изобелки и занимающими разные локусы на хромосоме, обнаружено большое число вариантов гемоглобина А, являющихся продуктами аллельных генов. 2. Группы крови Другой важный пример полиморфизма белков, связанный с проблемой переливания крови, — существование в популяции людей 3 аллельных вариантов гена фермента гликозил- трансферазы (А, В и 0). Этот фермент принимает участие в синтезе олигосахарида, локализованного на наружной поверхности плазматической мембраны и определяющего антигенные свойства эритроцитов. Варианты фермента А и В имеют разную субстратную специфичность: вариант А катализирует присоединение к олигосахариду N-ацетилгалактозамина, а вариант В — галактозы. Вариант 0 кодирует белок, лишённый ферментативной активности. В результате структура олигосахаридов, расположенных на поверхности эритроцитов, будет разной Антитела к антигенам А и В обычно имеются в сыворотке крови людей, на поверхности эритроцитов которых отсутствует соответствующий антиген, т. Е. индивидуумы с антигенами А на поверхности эритроцитов продуцируют в сыворотку крови антитела к В-антигенам (анти-В), а люди с В-антигенами — антитела к антигенам А (анти- А). В сыворотке крови анти-А и анти-В обычно присутствуют в высоких титрах и при появлении соответствующих антигенов способны активировать ферменты системы комплемента. При переливании крови руководствуются правилом, согласно которому кровь донора и реципиента не должна содержать антигены и антитела, реагирующие между собой: например, реципиенту, имеющему в сыворотке крови анти- А, нельзя переливать кровь от донора, содержащего на эритроцитах антигены А. При нарушении этого правила происходит реакция антиген-антитело. Это вызывает агглютинацию (склеивание) эритроцитов и их разрушение ферментами комплемента и фагоцитами. 3. Белки главного комплекса гистосовместимости и трансплантационная несовместимость При формировании клеточного иммунного ответа узнавание Т-лимфоцитами чужеродного антигена происходит только если он расположен рядом с гликопротеинами, присутствующими на собственной клеточной мембране. Эти гликопротеины называют белками главного комплекса гистосовместимости, или МНС-белками (см. раздел 1). Существуют 2 класса этих белков: молекулы класса I и II. МНС-белки класса I обнаружены практически во всех содержащих ядро клетках, включая Т- киллеры, тогда как МНС-белки класса II найдены главным образом в клетках, участвующих в иммунном ответе, в антиген-представляющих В- клетках и Т-хелперах, но не в Т-киллерах и макрофагах. Строение МНС-белков кодирует семейство генов, расположенных на коротком плече хромосомы 6 и занимающих участок ДНК длиной более 6000 пар нуклеотидов. Это семейство состоит из серии тесно сцепленных генов, ответственных за синтез МНС-белков и некоторых компонентов системы комплемента. Гены комплекса отличаются чрезвычайно высоким полиморфизмом. Число разных аллелей достигает нескольких миллионов. Белки МНС-системы считают самой полиморфной системой человека. Вариабельность МНС-белков обеспечивает трансплантационную несовместимость. Клетки трансплантата имеют набор этих белков, отличный от МНС-белков реципиента (во всех случаях, кроме генетически идентичных близнецов), и это приводит к развитию реакции клеточного иммунитета, в результате которой трансплантированная ткань отторгается. Исследования показали, что полиморфизм различных белков настолько велик, что можно говорить о биохимической индивидуальности и уникальности каждого человека. Глава 3.Наследственные болезни Каждый генетический локус характеризуется определённым уровнем изменчивости, т. Е. присутствием различных аллелей у разных индивидуумов. Аллели генов делят на 2 группы — нормальные, или аллели «дикого» типа, для которых функция гена не нарушена, и мутантные, приводящие к нарушению работы гена. «Плохой» аллель кодирует синтез белка, функция которого сильно нарушена и при гомозиготном наследовании фенотипически проявляется как наследственная болезнь. Наследственные болезни — следствие мутаций, произошедших в гаметах или зиготе. Такие мутации могут быть первичными, если возникли в гаметах или в процессе формирования зиготы, или вторичными, если мутантный ген возник раньше и был передан последующему поколению по наследству. Первичные мутации, как правило, не сопровождаются возникновением болезни, так как происходят обычно в одной из хромосом, и индивидуум, получивший такую мутацию, становится гетерозиготным носителем повреждения в гене. Мутантный ген в гетерозиготном состоянии часто не проявляется как болезнь и существенно не снижает жизнеспособность организма. Это способствует его распространению в популяции. При вторичных мутациях, если каждый из родителей является носителем мутантного гена, будучи гетерозиготой, возможно рождение детей-гомозигот по дефектному аллелю. В таком случае развивается наследственная болезнь, часто сопровождаемая очень тяжёлым течением. Согласно данным Всемирной организации здравоохранения, около 2,4% всех новорождённых на земном шаре страдают теми или иными наследственными нарушениями. Около 40% ранней младенческой смертности и инвалидности с детства обусловлены наследственной патологией. К настоящему времени на хромосомах человека выявлено около 800 генов, мутации в которых приводят к развитию различных наследственных болезней. Количество моногенных заболеваний (т.е. вызванных мутациями в определённом гене) ещё больше и равно примерно 950 в результате существования так называемых «аллельных серий», т. Е. групп болезней, клинически сильно отличающихся друг от друга, но обусловленных мутациями в одном и том же гене. Например, мутации в гене рецептора с тирозинкиназной активностью ret могут вызывать 4 различных наследственных заболевания. Более половины генов, в которых найдены мутации, вызывающие наследственные заболевания, охарактеризованы методами молекулярного анализа. Наибольшую по размеру группу составляют ферменты (31% от общего числа). За этой группой следуют белки, модулирующие функции белков и участвующие в правильном сворачивании полипептидных цепей (14%). На каждой хромосоме в среднем идентифицировано около 30 структурных генов, мутации в которых вызывают наследственные болезни. Однако распределены эти гены по хромосомам неравномерно. Так, например, на хромосоме 2 их в 3 раза меньше, чем на хромосоме 1. Наибольшее число мутантных генов (более 100) установлено на Х-хромосоме. Хорошо изученными наследственными заболеваниями, связанными с нарушением синтеза α- или β-цепей Нb, являются талассемии. Синтез α- и β-цепей в норме регулируется таким образом, что все молекулы протомеров используются на синтез тетрамера α2β2. Талассемии возникают как результат мутаций, включающих замены или делеции одного, или нескольких нуклеотидов, а иногда и целого гена, кодирующего структуру одного из протомеров. Эти болезни классифицируют по 4 типам: так, в случае, если одна из цепей не синтезируется, то их обозначают как α0- или β0-талассемии, а если синтез какой-либо из цепей снижен, то α+- или β+-талассемии. α-Талассемии возникают при нарушении синтеза α-цепей. В геноме каждого индивидуума существует 4 копии гена α-глобина (по 2 копии на каждой хромосоме), поэтому встречаются несколько видов недостаточности α-цепей. Если дефектна одна из 4 копий, то фенотипически это не проявляется, и такого человека рассматривают как «молчащего носителя» талассемии. При дефекте в 2 копиях гена у носителя мутации обнаруживают слабовыраженные признаки болезни, а при дефекте в 3 копиях развивается гемолитическая анемия. При полном отсутствии синтеза α-цепей (т. е. дефектны все 4 копии гена) наступает внутриутробная гибель плода. Β-Талассемии развиваются в результате снижения синтеза β-цепей Нb, для которых на каждой хромосоме имеется по одному гену. Синтез НbА начинается после рождения ребёнка. При дефекте в одной из копий гена недостаточность Нb проявляется в слабой степени и не требует специального лечения. Однако при полном выключении синтеза β-цепей развивается тяжёлая форма анемии, и таким пациентам проводят либо периодическую трансфузию крови, либо пересадку костного мозга. Со многими моногенными наследственными заболеваниями читатель познакомится практически во всех последующих разделах учебника. Здесь же хотелось бы только отметить, что наряду с болезнями, наследственная природа которых ярко выражена, существует множество болезней, характеризующихся семейной предрасположенностью. Это такие широко распространённые заболевания, как сахарный диабет, подагра, атеросклероз, шизофрения и ряд других. В отличие от моногенных болезней, эти заболевания относят к мультифакторным. Поэтому исследования, направленные на выявление белков, аллельные формы которых ответственный за предрасположенность к заболеванию, являются задачами настоящего и будущего времени. Заключение В настоящее время стало очевидным, что достижения в области молекулярной биологии способны сильно изменить практическую медицину. Они не только углубили наши знания об экспрессии генов и причинах многих болезней, но способствовали разработке новых подходов к их диагностике и лечению. Было установлено, что полиморфизм генов широко распространён в популяции людей, показана взаимосвязь между изменениями в структуре ДНК и многими болезнями. Идентификация генов, нарушение работы которых приводит к развитию наследственных заболеваний, создала предпосылки для подробного анализа генетических и биохимических основ патогенеза этих заболеваний и разработки наиболее эффективных методов лечения. Методами молекулярной медицины были созданы вакцины для предотвращения гепатитов, инсулин человека — для лечения сахарного диабета, фактор VIII — для восстановления нормального свёртывания крови и лечения гемофилии и многие другие препараты. С помощью генной терапии оказалось возможным вводить в организм больного полноценно работающие гены и таким образом восстанавливать метаболические нарушения, вызванные мутантными генами. Список литературы 1.БИОХИМИЯ УЧЕБНИК ДЛЯ ВУЗОВ – Е. С. Северина – 2004 стр. 199-211 2. https://lifelib.info/biochemistry/biochemistry_4/27.html 3. http://www.cnshb.ru/AKDiL/0037t/base/RP/000653.shtm |