генетика бактерий. Методы изучения генетики бактерий
 Скачать 77.68 Kb. Скачать 77.68 Kb.
|

5 по 0,1мл высевают на плотную селективную питательную среду (среда Эндо) в чашки Петри и растирают шпателем.Тема Методы изучения генетики бактерий Содержание Введение…………………………………………………………………………3 Глава 1: Современное представление о генетическом аппарате бактерий…..5 Составляющие элементы генетического аппарата бактерий Особенности генетического материала бактерий Способы переноса наследственного материала у бактерий Влияние внешних факторов на функционирование генетической системы бактерий Глава 2: Современные методы изучения генетики бактерий………………....14 2.1 Методы выявления фенотипические изменчивости………………………14 2.2 Методы………………………………………………………………………..15 Метод определения Col-плазмид Метод определения колицинотипа Метод выявления спонтанности мутаций Метод индукции мутаций под действием ультрафиолетового излучения 2.3 Методы определения……………………………………………………..…21 Метод постановки опыта конъюгации Метод постановки опыта трансформации Метод постановки опыта специфической трансдукции Перспективы Заключение……………………………………………………………………...26 Список использованной литературы…………………………………………..27 Введение Человек использовал бактерии, ещё не зная об их существовании. С помощью заквасок, содержащих Бактерии, приготовляли кисломолочные продукты, уксус, тесто и т. д. Впервые бактерии увидел А. Левенгук -- создатель микроскопа, исследуя растительные настои и зубной налёт. К концу 19 -- началу 20 вв. было выделено большое число бактерий, обитающих в почве, воде, пищевых продуктах и т. п., были открыты многие виды болезнетворных бактерий. Физиология микроорганизмов изучает жизнедеятельность микробных клеток, процессы их питания, дыхания, роста, размножения, закономерности взаимодействия с окружающей средой. Выяснение физиологии этих микроорганизмов важно для постановки микробиологического диагноза, проведения лечения и профилактики инфекционных заболеваний, регуляции взаимоотношений животного организма с окружающей средой. Генетика (от греческого genesis - происхождение) - наука о наследственности и изменчивости организмов. Начало генетики как науки положил Мендель, который, скрещивая между собой сорта гороха с качественно различающимися признаками, установил главные закономерности наследственности. Изучение и анализ законов передачи наследственных признаков от поколения к поколению, а также выяснение механизмов, обеспечивающих наследования на всех уровнях организации живых существ, является главной целью генетики. Успехи генетики стали возможны благодаря совместной работе микробиологов, генетиков, химиков и физиков, которые в своих исследованиях использовали микроорганизмы. Именно бактерии и вирусы оказались наиболее простой и удобной моделью для решения насущных проблем молекулярной генетики, изучение которых и стало актуальностью данной работы. Цель работы – рассмотреть современные методы изучения генетики бактерий. Для решения поставленной цели необходимо решить следующие задачи: - изучить оставляющие элементы генетического аппарата бактерий; - рассмотреть влияние внешних факторов на функционирование генетической системы бактерий; - дать характеристику современным методам изучения генетики бактерий; Объектом данной работы является генетика бактерий. Предмет работы - методы изучения генетики бактерий. Структура работы состоит из введения, двух глав, заключения и списка использованной литературы. Глава 1. Современное представление о генетическом аппарате бактерий Составляющие элементы генетического аппарата бактерий Генетический аппарат бактерий представлен хромосомными (нуклеоид) и внехромосомными (плазмиды, инсерционные последовательности, транспозоны) структурами. Нуклеоид: - не имеет ядерной мембраны и не связан с гистонами; - одна непарная суперспирализованная хромосома состоит из двунитевой молекулы ДНК (кольцевой или линейной) размером от 3х108 до 2,9х109 Д и содержит до 4600 генов; один конец бактериальной хромосомы связан с мезосомой; некоторые бактерии имеют сложные геномы, состоящие из двух или нескольких репликонов. Структура ДНК и генетический код.Материальной основой наследственности, определяющей генетические свойства всех организмов, является ДНК. Исключение составляют только РНК–содержащие вирусы, у которых генетическая информация закодирована в РНК [8]. ДНКсостоит из последовательности химически связанных нуклеотидов и имеет структуру правильной двойной спирали из закрученных одна вокруг другой двух полинуклеотидных цепей. Каждый нуклеотид состоит из азотистого основания, сахара дезоксирибозы и фосфатной группы. Азотистые основания представлены пуринами (аденин, гуанин) и пиримидинами (тимин, цитозин). ДНК содержит А, С, G, Т; РНК— А, С, G, U. Каждый нуклеотид обладает полярностью. У него имеются дезоксирибозный 3'–конец и фосфатный 5'–конец. Нуклеотиды соединяются в полинуклеотидную цепочку фосфодиэфирными связями между 5'–концом одного нуклеотида и 3'–концом другого. Соединение между двумя цепочками обеспечивается водородными связями комплементарных азотистых оснований: аденина с тимином, гуанина с цитозином. Нуклеотидные цепи антипараллельны: на каждом конце линейной молекулы ДНК расположены 5'–конец одной цепи и 3'–конец другой цепи. Последовательность нуклеотидов ДНК определяет последовательность аминокислотных остатков в молекуле белка. Каждому белку соответствует свой ген — уникальная структурная единица наследственности. Ген — фрагмент полинуклеотидной цепи молекулы ДНК, отличающийся числом и специфичностью последовательности нуклеотидов и кодирующий синтез одного пептида. В ДНК содержатся структурные и регуляторные гены. Структурные гены несут информацию о синтезируемых ферментах или структурных белках. Гены, ответственные за синтез вещества, обозначают строчными буквами латинского алфавита, соответствующими названию данного вещества со знаком «+» (his+— гистидиновый ген, leu+ — лейциновый ген). Гены, контролирующие резистентность к лекарственным препаратам, фагам, обозначают буквой r (resistent— резистентный). Напр., резистентность к стрептомицину записывается strr, а чувствительность — strs. Регуляторные гены регулируют транскрипцию структурных генов. Хромосома состоит из особых функциональных единиц — оперонов. Оперон — совокупность промотора, оператора и структурных генов — является функциональной генетической единицей, регулирующей экспрессию одного или группы генов. Основные этапы развития генетической системы: кодон ген оперон геном вирусов и плазмид хромосома прокариот (нуклеоид) хромосомы эукариот (ядро). Регуляция выражения генетической информации у бактерий. Бактериальная клетка способна запустить или прекратить синтез фермента в зависимости от присутствия соответствующего субстрата. Для этого бактериальные гены объединены в группы так, что все ферменты, необходимые для осуществления биосинтеза, детерминируются генами, сцепленными друг с другом. Вся группа генов может транскрибироваться в одну полицистронную мРНК, которая последовательно транслируется рибосомами с образованием каждого из белков. Экспрессия генов у прокариот регулируется на уровне транскрипции. Роль сигнальных веществ для запуска транскрипции играют низкомолекулярные соединения, которые являются либо субстратом для фермента, либо продуктом ферментативной деятельности. Индукция и репрессия представляют собой разные стороны одного и того же явления. Малые молекулы, индуцирующие образование ферментов, способных метаболизировать их, называются индукторами. Те же, которые предотвращают образование ферментов, способных синтезировать их, —корепрессорами [5]. Молекулы-эффекторы не могут вступать в прямое взаимодействие с ДНК, посредником для них служит специальный регуляторный белок. Регуляторный белок, который связывается с ДНК в отсутствии индуктора, называется репрессором. За синтез регуляторных белков ответственны регуляторные гены. В присутствии белка-репрессора транскрипция блокирована; его удаление обусловливает доступ РНК-полимеразы к генам и запуск транскрипции. Прекращение синтеза фермента при помощи белка-репрессора получило название репрессии. Репрессия позволяет бактериальной клетке избежать перевода своих ресурсов на ненужную в данный момент синтетическую активность. Если индуктор присутствует в клетке в высокой концентрации, то в результате специфического присоединения к регуляторному белку он изменяет его конформацию и способность связываться с ДНК. Контроль транскрипции достигается взаимодействием регуляторного белка с регуляторным сайтом (оператором), который расположен между структурными генами и промотором (участком, распознаваемым ДНК-зависимой РНК-полимеразой). Промотор служит местом связывания РНК-полимеразы, и от него начинается транскрипция. Особенности генетического материала бактерий Генетическая система бактерий имеет 4 особенности: 1.Хромосомы бактерий и плазмид располагаются свободно в цитоплазме неограниченной от неё мембранами. Так как длина хромосомы во много раз превышает длину бактериальной клетки, то хромосома компактным образом в ней упакована, молекула ДНК находится в суперсперализированной форме. Такая упаковка способствует биосинтезу белка. 2.Бактерии являются гаплоидными организмами – имеют один набор генов. Содержание ДНК не постоянно, может не в благоприятных условиях достигать значений эквивалентных по массе 2,4,6 и 8 хромосомами. У всех других живых существ ДНК постоянная и она удваивается (кроме вирусов и плазмид) перед делением. 3.У бактерий в естественных условиях передача генетической информации происходит не только по вертикали, т.е. от родительской клетки к дочерним, но и по горизонтали, с помощью механизма конъюгации, трансдукции, трансформации и другие. 4.У бактерий кроме хромосомного генома есть дополнительный - плазмидный геном, наделяющий бактерию-хозяина, важными биологическими свойствами: специфическим иммунитетом к различным антибиотика и другим химическим препаратам. Если у бактерии увеличивается содержание ДНК, то также увеличивается количество рибосом. Содержание ДНК у бактерий зависит от условий роста. В благоприятных условиях оно возрастает. Регулируя содержание своих генов, бактерии приспосабливают скорость размножения к условиям роста. Возможность регулировать свое размножение является главным условием выживания бактерий в окружающей среде и сохранения вида в природе. Способы переноса наследственного материала у бактерий Быстрая реакция на изменения условий окружающей среды и приобретение необходимых генетических признаков (напр. изменение вирулентных свойств, устойчивость к АМП). Обмен генетической информацией (ОГИ)- механизм, обеспечивающий многообразие микробного мира [6]. ОГИ способствует быстрой адаптации бактерий. ОГИ приводит к образованию рекомбинантной ДНК. ОГИ служит важным фактором эволюции бактерий. 3 способа передачи генетической информации: 1. Конъюгация – внедрение ДНК из бактерии – донора в клетку реципиента 2. Трансформация – поглощение свободной ДНК из внешней среды 3. Трансдукция – распространение генов бактерий умеренными фагами Схема конъюгации Конъюгация – однонаправленный перенос генетического материала (хромосомной и плазмидной ДНК) от донора к реципиенту при непосредственном контакте клеток. Донор – бактерия, содержащая конъюгативную плазмиду или конъюгативный транспозон, способная передавать ДНК реципиенту путем горизонтального переноса генов. Реципиент – любая бактериальная клетка, способная принимать ДНК от бактерии-донора путем горизонтального переноса генов. Механизм передачи конъюгативных плазмид между Гр- бактериями: 1. Образование межклеточного контакта - конъюгативных пилей 2.Сайт-специфическое разрезание одной цепочки ДНК 3.Сборка особой структуры - релаксосомы для переноса ДНК 4.Перемещение разделенных цепей ДНК в клетку реципиента 5.Синтез комплементарных цепей ДНК 6. Терминация переноса ДНК и разделение конъюгирующих клеток Трансформация -поглощение бактерией ДНК из окружающей среды и включение в свой геном, при этом бактерия становится генетически трансформированной. Впервые этот феномен описан у Streptococcus pneumoniae, когда непатогенные живые мутантные клетки стали патогенными для мышей при одновременной инъекции с убитыми нагреванием вирулентными клетками S.pneumoniae. Изменение происходило в результате трансформирующего действия ДНК из убитых нагреванием вирулентных клеток S.pneumoniae. Схема трансформации 1. Адсорбция ДНК на клетке (клеточной стенке) 2. Проникновение внутрь клетки 3. Рекомбинация с ДНК клеточной хромосомы Трансдукция - обмен генетическим материалом с участием бактериофагов. Бактериофаги– вирусы бактерий - молекулярные паразиты, использующие для самовоспроизводства аппарат репликации ДНК и синтеза белка зараженных ими клеток. Трансдукцию осуществляют бактериофаги, содержащие невирусную генетическую информацию. Подобно плазмидам, бактериофаги могут выступать в качестве векторов переноса ДНК от клеток-хозяев. Перенос фаговой информации может служить механизмом обмена генетическим материалом между бактериями. Бактериофаги E.coli Виды трансдукции: 1. Специфическая трансдукция – перенос генов клетки-хозяина из строго определенных участков хромосомы бактериальной клетки. (Например, 5 из 11 фагов возбудителя дифтерии Corynebacterium diphtheriae – переносчики генов дифтерийного токсина). 2.Неспецифическая трансдукция (общая) - случайный перенос различных генов клетки-хозяина, расположенных в разных участках генома бактерии.. Схема трансдукции 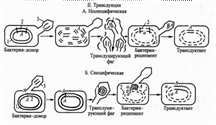 Влияние внешних факторов на функционирование генетической системы бактерий. Жизнедеятельность микробов находится в зависимости от окружающей среды. Создавая те или иные условия в среде, где развиваются микробы, можно способствовать развитию полезных и подавлять жизнедеятельность вредных микроорганизмов. Пищевые продукты могут хорошо сохраняться только при создании неблагоприятных условий для развития в них вредных микробов. Основными факторами, влияющими на жизнедеятельность микробов, являются: температура, влажность, действие света, характер питательной среды [7]. Температура. Все микробы имеют максимальную, оптимальную и минимальную температуру своего развития. Оптимальная температура для большинства микроорганизмов 25—35°С. Поэтому пищевые продукты в этих условиях быстро портятся. Минимальный температурный предел у разных микробов различен. Понижение температуры замедляет или прекращает развитие микробов, но не убивает их. Поэтому при охлаждении (6°С) и замораживании (от -6 до -20°С) пищевые продукты хорошо сохраняются, но при оттаивании и обработке их микробы вновь начинают свою деятельность. Максимальная температура (45—50°С) также приостанавливает развитие микробов. Дальнейшее повышение температуры ведет к гибели вегетативных клеток, а затем и спор. На губительном действии высоких температур на микробы основаны пастеризация (60—90°С) и стерилизация (100— 120°С) пищевых продуктов. В зависимости от температуры развития микробы делят на психро-филъные(холодоустойчивые), у которых оптимум развития 15°С (плесневые грибы); мезофилъные(развивающиеся при средней температуре), у которых оптимум 25—37° С (болезнетворные бактерии, дрожжи); термофильные (теплолюбивые), у которых оптимум 50° С (молочно-кислые бактерии). Влажность. Повышенная влажность увеличивает количество растворимых питательных веществ, следовательно, способствует питанию и развитию микробов. Поэтому пищевые продукты, содержащие большое количество влаги (молоко, мясо, рыба, овощи, плоды), быстро портятся. Нижний предел влажности среды для развития бактерий 20 %, а плесневых грибов — 15 %. Поэтому надежным способом сохранения продуктов от порчи является их сушка до влажности ниже указанного предела. Среда с повышенной концентрацией веществ. Как уже было сказано, микробы живут в среде с небольшой концентрацией растворимых веществ. При повышении концентрации соли (до 10—20 %) и сахара (до 60—70 %) многие микробы полностью прекращают свое развитие (гнилостные, молочнокислые) в результате обезвоживания микробных клеток. Действие высокой концентрации соли на микробы используют при посоле рыбы, мяса, а сахара — при приготовлении варенья, джема, повидла. Реакция среды. Большинство микробов развивается в нейтральной (рН = 7) или слабощелочной среде (рН = 8), а плесени и дрожжи — в слабокислой среде (рН = 3-6). Изменяя реакцию среды, можно влиять на ход развития микроорганизмов. На этом основаны способы консервирования овощей, плодов путем квашения (с помощью образующейся молочной кислоты) и маринования (с помощью добавляемой уксусной кислоты), в процессе которых подавляется жизнедеятельность гнилостных микробов. Свет. Прямой солнечный луч губит микробы, в том числе и болезнетворные. Губительны ультрафиолетовые лучи солнца и специальных ламп БУВ, используемых для дезинфекции воды, воздуха, помещений. Химические вещества. Многие химические соединения губительно действуют на микробов и используются для их уничтожения. Они называются антисептиками или дезинфицирующими веществами. Так, хлорную известь в общественном питании применяют для дезинфекции рук, посуды и оборудования (0,2 %), сорбиновуюкислоту—для сохранения соков. Наличие бензойной кислоты в клюкве, бруснике предохраняет их от порчи [10]. Биологические факторы. Микробы в процессе жизнедеятельности могут влиять друг на друга, способствуя развитию или угнетению. Последнее свойство микробов используется человеком в борьбе с болезнетворными микробами. Многие бактерии, плесневые грибы выделяют в окружающую среду вещества — антибиотики, губительно действующие на развитие других микробов. Пенициллин, стрептомицин, грамицидин, биомицин — антибиотики, широко применяемые в медицине. Другими веществами, близкими к антибиотикам по характеру действия на микробов, являются фитонциды. Эти вещества, выделяемые многими растениями (луком, чесноком, хреном, цитрусовыми и др.), убивают болезнетворные микробы дизентерии, гнилостную палочку и др. Глава 2. Современные методы изучения генетики бактерий 2.1 Методы выявления фенотипические изменчивости Модификация микроорганизмов возникает как ответ клетки на неблагоприятные условия ее существования. Это адаптивная реакция на внешние раздражители. Модификация не сопровождается изменением генотипа, в связи с чем возникшие в клетке изменения по наследству не передаются. При восстановлении оптимальных условий возникшие изменения утрачиваются. Модификация может касаться разных свойств микроорганизмов -- морфологических, культуральных, биохимических и др. Морфологическая модификация выражается в изменениях формы и величины бактерий. Например, при добавлении пенициллина к питательной среде клетки некоторых бактерий удлиняются. Недостаток в среде солей кальция вызывает у палочки сибирской язвы повышенное спорообразование. При повышенной концентрации солей кальция способность образовывать споры утрачивается и т. д. При длительном росте бактерий в одной и той же среде возникает полиморфизм, обусловленный влиянием накопившихся в ней продуктов их жизнедеятельности. Культуральная модификация состоит в изменении культуральных свойств бактерий при изменении состава питательной среды. Например, при недостатке кислорода у стафилококка утрачивается способность образовывать пигмент. Чудесная палочка при комнатной температуре образует ярко-красный пигмент, но при 37 °С способность образовывать этот пигмент утрачивается и т. д. Биохимическая (ферментативная) модификация. Каждый вид бактерий имеет определенный набор ферментов, благодаря которым они усваивают питательные вещества. Эти ферменты вырабатываются на определенных питательных субстратах и предопределены генотипом [9]. В процессе жизнедеятельности бактерий обычно функционируют не все гены, ответственные за синтез соответствующих ферментов. В геноме бактерий всегда имеются запасные возможности, т.е. гены, определяющие выработку адаптивных ферментов. Например, кишечная палочка, растущая на среде, не содержащей углевод лактозу, не вырабатывает фермент лактазу, но если пересеять ее на среду с лактозой, то она начинает вырабатывать этот фермент. Адаптивные ферменты позволяют приспособляться к определенным условиям существования. Таким образом, модификация — это способ приспособления микроорганизма к условиям внешней среды, обеспечивающий им возможность расти и размножаться в измененных условиях. Приобретенные свойства не передаются по наследству, поэтому они не играют роли в эволюции, а способствуют в основном выживанию микробных популяций. 2.2 Методы Метод определения Col-плазмид Col-плазмиды, Col-факторы, или факторы колициногенности определяют способность бактериальных клеток штаммов Е.соК синтезировать белки - колицины, вызывающие гибель клеток, не имеющих иммунитета к колицину данного типа. Col-плазмиды подразделяют на 2 группы. К первой группе относят плазмиды: ColEl, Со1Е2, Со1ЕЗ, ColE4, ColE5, C0IE6, ColE7, C0IE8, ColE9, ColN, ColK, ColA, ко второй - Collb, ColB, ColV. Феномен колициногении был открыт A. Gratia в 1925 году у бактерий штамма E.coli. Как оказалось позднее, плазмиды колициногении достаточно широко распространены и отличаются размерами, копий- ностью, принадлежностью к разным группам несовместимости. Важно также упомянуть, что не все гены колицинов являются плазмидными, гены колицинов L и микроцина Е 492 - нуклеоидные. Одна бактериальная клетка может нести несколько разных Col - плазмид. Из чего можно сделать вывод о принадлежности указанных плазмид к разным группам несовместимоти и, следовательно, отсутствии филогенетического родства между ними [7]. Среди Col - плазмид встречаются как конъюгативные (Collb, ColB и др.), так и неконъюгативные (ColEl, Со1Е2 и др.) плазмиды. Конъюгативные плазмиды присутствуют в клетке в 2–3 копиях и имеют размеры более 60 МД, а некоторые (ColV) могут интегрироваться в нуклеоид, формируя клетки Hfr. Такие плазмиды не только сами передаются в другие клетки, но и способны мобилизовать нуклеоид. Неконъюгативные Col-плазмиды (ColEl, Со1Е2 и др.) имеют меньшие размеры (приблизительно 5 МД) и являются многокопийны- ми [7]. В настоящее время известно более 20 различных типов колицинов, характеризующихся довольно сложными механизмами синтеза и разными механизмами действия. Показано, что токсин, адсорбируясь на поверхности клетки, изменяет процессы ее метаболизма, провоцируя гибель. В плане воздействия на клетку можно упомянуть следующее: токсин Е1 изменяет цитолемму бактериальной клетки, колицин ЕЗ оказывает воздействие на рибосомальную РНК (16SpPHK). Для секреции колицина ColV необходим специальный экспортер, включающий плазмидные белки CvaA, CvaB и TolC, при этом, основная роль в процессинге ColV отводится белкам CvaA и CvaB. Кроме того, было показано, что эффективность работы генов, синтезирующих колицин, и количество колицина в клетке возрастает после повреждения ДНК клетки-хозяина. Существуют плазмидные гены, ответственные за выход колицинов из клетки в окружающую среду. Нук- леоидные гены участвуют в регуляции экспрессии колициновых генов. Как показано, белок гена texA репрессирует транскрипцию гена коли- цина Coll [265]. Интересен факт вовлечения в синтез токсина только малого числа плазмидосодержащих клеток бактериальной популяции до воздействия деструктивных агентов. Появление повреждений приводит к резкому повышению уровня синтеза колицина, что походит на формирование sos-ответа, который нуждается в белках генов гесА, 1ехА [87]. Иммунитет клетки-хозяина, несущей определенную Col- плазмиду, обусловлен одновременным синтезом соответствующего колицина и иммунного белка. У одних плазмид гены двух указанных белков могут быть локализованными в одном опероне плазмидного генома, тогда как у других - в разных [2]. Таким образом, Col-плазмиды обеспечивают бактериальные клетки определенными преимуществами при заселении новых экологических ниш. Метод определения колицинотипа Определение спектра чувствительности культур Зонне к колицинам проводят при помощи эталонных колициногенных штаммов Фредерика:
Колицинотипирование, т. е. определение спектра чувствительности культур Зонне к колицинам, определяют следующим образом. Свежие эталонные колициногенные штаммы Фредерика, выросшие на среде Ресселя, сеют уколом петли в соответствующие квадраты чашки с 1,5%-ным МПА, подкрашенным генцианвиолетом, и инкубируют при 37ºC в течение 24 часов. Выросшие колонии бактерии убивают парами хлороформа, и поверхность агара заливают 2,5 мл расплавленного 0,7%-ного МПА, содержащего 2 капли 6-часовой испытуемой бульонной культуры шигелл Зонне. После 20-часового роста при 37°C производят учет результатов опыта. При наличии чувствительности к колицинам испытуемого штамма бактерии Зонне вокруг колоний колициногенных культур образуются зоны задержки роста подобно зонам задержки роста вокруг дисков с антибиотиками. Культура считается чувствительной к колицину, если ширина задержки роста бактерий более 2 мм [3]. Результаты регистрируют плюсами: + - зона задержки 2 мм; ++ - зона задержки 2-5 мм; +++ - зона задержки 5-10 мм; ++++ - зона задержки более 10 мм. Б. Вирулентность дизентерийных бактерий обусловлена наличием у них факторов адгезии и колонизации, инвазии. (Эти факторы патогенности контролируются главным образом плазмидами). Кроме того, шигеллы способны вырабатывать 2 типа экзотоксинов: цитотонины (термолабильный и термостабильный) и цитотоксины (способностью вырабатывать цитотоксины шигеллу наделяют конвертирующие умеренные фаги). Патогенность шигелл определяется с помощью кератоконъюнктивальной пробы на морской свинке. В. Факторы патогенности диареегенных e. Coli
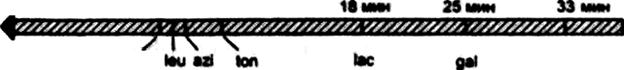
٭ - имеются в виду CFA/I, CFA/II, CFA/III, CFA/IV. Метод выявления спонтанности мутаций В две чашки Петри с питательным агаром вносят по 0,1 мл суточной культуры E.coli М17 и распределяют равномерно по поверхности питательной среды. Через 6 ч инкубации при 370С в одной из чашек перераспределяют шпателем выросшие микроколонии. Через 24 ч из каждой чашки культуры пересевают методом отпечатков на поверхность питательного агара с рифампицином. Через 24 ч инкубации учитывают результат: на поверхности среды с рифампицином в чашке без перераспределения выросли единичные колонии рифампицинрезистентных мутантов, а в чашке с перераспределением выросли более многочисленные (в десятки – сотни раз) колонии антибиотикоустойчивых мутантов. Данный опыт показывает, что антибиотикоустойчивые мутанты возникли спонтанно, до контакта бактерий с селективным агентом – рифампицином. Уже через 6 ч на среде без антибиотика появляются микроколонии антибиотикоустойчивых мутантов. Благодаря перераспределению бактерии мутанты из этих микроколоний распространяются по всей поверхности среды и после посева отпечатками на среду с антибиотиками дадут начало многочисленным колониям мутантов, в то время как отпечатки с чашки без перераспределения выявляют только небольшое число колоний, соответственно исходным микроколониям мутантов. Метод индукции мутаций под действием ультрафиолетового излучения Воздействие УФИ индуцирует у бактерий, в частности Е. coli, предмутационные повреждения и широкий спектр мутаций, включая замены оснований, образование делений и др. В качестве источника УФИ используют бактерицидную лампу ВУФ-15, которую устанавливают на расстоянии 60 см от центра облучаемого объекта. Поскольку Е. coli обладает эффективными механизмами световой репарации УФ-поврсждений ДНК, то облучение желательно проводить в темноте или при красном свете. Предварительно определяют бактерицидную дозу УФ-облучения, на основании которой устанавливают оптимальную мутагенную дозу, равную 0,1... 1 % от числа выживших бактерий [6]. Для получения lac-мутантов Е. coli испытуемую культуру предварительно выращивают в питательном бульоне в течение 14...18 ч. При этом концентрация бактериальных клеток в 1 мл среды составляет примерно 2 ■ 108. Клетки осаждают центрифугированием, ресуспендируют в 40 мл 0,1 М раствора MgSO4 и охлаждают во льду для прекращения деления клеток. Суспензию в объеме 50 мл помещают в стерильную чашку Петри и облучают в течение 15... 150 с, после чего клетки осаждают центрифугированием, ресуспендируют в таком же объеме питательного бульона. Пробирку с бульонной культурой инкубируют при 37 "С в течение 14...18ч. Затем из разведений 10 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Свойства | E. coli | | |
| донор (F+,leu+, strs) | реципиент (F-, leu-, strr) | рекомбинант (F+, leu+, strr) | |
| Наличие F-плазмиды | да | нет | да |
| Синтез лейцина | да | нет | да |
| Устойчивость к стрептомицину | нет | да | да |
| Рост на минимальной среде (без лейцина) со стрептомицином | нет | нет | да |
Метод постановки опыта трансформации
Трансформация - горизонтальный перенос генов от донора к реципиенту через внешнюю среду; осуществляемый после гибели бактерий-доноров.
Для проведения опыта необходимы следующие материалы:
-реципиент - штамм Bacillus subtilis (strs), чувствительный к стрептомицину;
- ДНК донора – выделяют из штамма B. subtilis (strr), устойчивого к стрептомицину;
- селективная среда - МПА со стрептомицином.
Постановка опыта трансформации. Смешивают по 1мл бульонной культуры B. subtilis и ДНК донора. Инкубируют 30 мин при +370С, и высевают на МПА и МПА со стрептомицином. Рекомбинантные штаммы способны расти на селективной среде со стрептомицином. Частота трансформаций равна отношению количества рекомбинантных клеток к реципиентным.
Метод постановки опыта специфической трансдукции
Трансдукция– горизонтальный перенос генов от донора к реципиенту умеренными бактериофагами.
Для доказательства существования трансдукции может быть приведен опыт по горизонтальному переносу от донора к реципиенту геновβ-галактозидазного оперона, контролирующего расщепление лактозы у Е. coli. Для проведения опыта необходимы:
-реципиент - штамм E. coli, лишенный β-галактозидазного оперона (E. coli lac-), на среде Эндо образует бесцветные колонии;
- трансдуцирующий фаг - фаг (λ dgal), в геноме которого часть генов замещена генами β-галактозидазного оперона E. coli. Концентрация фага - 106–107 частиц в 1 мл;
- селективная среда Эндо, на которой лактозоотрицательные колонии бесцветны, а лактозоположительные колонии - ярко малиновые, с металлическим оттенком.
Постановка опыта трансдукции. Смешивают по 1 мл трехчасовой бульонной культуры реципиента и трансдуцирующего фага. Смесь инкубируют 60 мин при +370С, готовят серию десятикратных разведений. Из пробирки с разведением 10-6 по 0,1 мл культуры высевают на среду Эндо и инкубируют в течение суток. Рекомбинантные клетки растут с образованием малиновых с металлическим оттенком колоний. Частота трансдукций равна отношению количества клеток рекомбинантов к числу реципиентов.
Перспективы
Получение наследственно измененных форм микроорганизмов расширило возможности их использования в сельскохозяйственном и промышленном производстве, а также в медицине. Основной метод получения новых форм микроорганизмов — индуцирование мутаций воздействием различными мутагенами на дикие, существующие в природе культуры. Таким методом удается создавать мутантов, которые выделяют в десятки и сотни раз большее количество ценных продуктов (антибиотиков, ферментов, витаминов, аминокислот и т. д.) по сравнению с дикими формами микроорганизмов.
Процесс получения высокопродуктивных штаммов микроорганизмов состоит из многих этапов. Сначала на культуру микроорганизма воздействуют различными мутагенными факторами с последующим отбором наиболее продуктивного штамма. Мутантный штамм могут подвергнуть дальнейшему воздействию мутагенов и последующему отбору еще более продуктивных форм. Часто из тысяч бесполезных мутантов отбирают только один высокопродуктивный штамм. В последние годы методом радиационного и химического мутагенеза микроорганизмов получено большое число промышленных штаммов микроорганизмов — продуцентов нужных человеку веществ [7].
Особенно широкие перспективы переделки наследственной природы организмов сулит развитие генной, или генетической, инженерии — раздела молекулярной генетики, разрабатывающего методы создания новых генетических структур с заданной информацией и способы их переноса в клетки прокариот и эукариот.
Полученные методом генной инженерии новые генетические молекулы представляют собой рекомбинантные ДНК, включающие два компонента — вектор (переносчик) и клонируемую «чужеродную» ДНК. Поскольку переносчик должен обладать свойствами репликона и обусловливать репликацию вновь созданной реком-бинантной ДНК, то вектором обычно служат такие репликоны, как плазмиды, умеренные фаги и вирусы животных. Все упомянутые переносчики имеют циркулярно замкнутую структуру ДНК. Клонируемая ДНК — это фрагмент ДНК, несущий необходимый ген (или гены), контролирующий образование нужного вещества.
Существуют различные приемы получения рекомбинантных молекул ДНК. Наиболее простой из них начинается с обработки изолированных молекул ДНК-вектора и ДНК, несущей необходимый ген, ферментами-рестриктазами (эндонуклеазы рестрикции), расщепляющими взятые молекулы ДНК в строго определенном месте с образованием однонитчатых, комплементарных друг другу концов, так называемых «липких концов». Таков первый этап получения рекомбинантных ДНК, его иначе называют «разрезание» молекул ДНК при помощи эндонуклеаз рестрикции. Второй этап заключается в обработке полученных линейных молекул ДНК ферментом полинуклеотидлигазой, которая «сшивает» две разные молекулы в одну рекомбинантную ДНК. На третьем этапе рекомбинантные молекулы вводят в клетки тех или иных бактерий (методом трансформации. На завершающем, четверто, этапе выполняют клонирование трансформированных клеток.
Методом генной инженерии уже получены рекомбинантные молекулы ДНК, несущие информацию для образования таких важных веществ, как интерферон, инсулин, гормон роста человека и другие, в клетках кишечной палочки. По-видимому, тем же метолом можно будет создать и такие бактерии, которые, потеряв свою болезнетворность, помогут выработать иммунитет против многих инфекционных болезней животных и человека. В промышленности благодаря использованию генной инженерии появятся высокопродуктивные микроорганизмы, создающие белки, ферменты, витамины, антибиотики, ростовые вещества и другие нужные продукты.
Возможно, будут получены новые сорта растений и породы животных, устойчивые к заболеваниям и в полной мере наделенные хозяйственно ценными свойствами. Метод генной инженерии поможет вывести формы растений, обладающие способностью к связыванию молекулярного азота атмосферы. Такие растения, вероятно, можно будет получить после введения в их геном генов от микроорганизмов, фиксирующих азот из воздуха [10].
В связи с разработкой и совершенствованием методов генной инженерии, показавших возможность передачи не только естественных генов живых организмов, но и искусственно синтезированных, открываются блестящие перспективы для научно-технического прогресса не только в медицине и промышленности, но и в сельскохозяйственном производстве.
Заключение
Генетика микроорганизмов изучает наследуемые изменения свойств микробов, возникающие самопроизвольно (спонтанные мутации) или в результате различных химических и физических воздействий (индуцированные мутации), а также процессы обмена генетическим материалом между микроорганизмами, структуру и функцию их генетического аппарата.
Обмен генетическим материалом у бактерий осуществляется тремя различными способами:
1) трансформация бактерий; при этом в бактерию-реципиент вносится часть генов бактерии-донора в виде изолированной молекулы ДНК;
2) трансдукция бактерий; в этом случае роль переносчика генетического материала между клетками донора и реципиента выполняют умеренные бактериофаги. В первом и втором способах не требуется непосредственного контакта между донором и реципиентом;
3) конъюгация бактерий; при этом обмен генетическим материалом происходит в момент непосредственного контакта между донором и реципиентом.
После попадания генетического материала донора в бактерию-реципиент происходит собственно генетический обмен: рекомбинация между молекулами ДНК. Генетический обмен у вирусов осуществляется при совместном размножении двух или более вирусных частиц внутри одной клетки.
Успехи современной генетики микроорганизмов позволили объяснить ряд явлений и процессов, важных для практики. Выяснены механизмы формирования лекарственной устойчивости бактерий и намечены пути для ее устранения. Получены мутанты — активные продуценты антибиотиков, витаминов и аминокислот, важных для медицинской и хозяйственной практики.
Список использованной литературы
1. П.О.Кимович Медицинская микробиология под редакцией акад. РАМН В.И.Покровского. -М.2016г.
2. В.Д.Тимаков, В.С. Левашев, Л.Б.Борисов Микробиология: Учебник.-2-е изд.,перераб.-М.,2013, 512с.
3. Поздеев О.К. Медицинская микробиология. 2019г.
4. http://032.help-rus-student.ru/text/002.htm
5. http://www.o-med.ru/083.php
6. Земсков М. В. и др Основы общей микробиологии, вирусологии и иммунологии. Изд. 2-е, испр. и доп. М., «Колос», 2017.
7. http://collegemicrob.narod.ru/microbilogy/
8. Жимулёв И.Ф. Общая и молекулярная генетика: учебное пособие для студентов университетов, обучающихся по направлению 510600 - Биология и биологическим специальностям. - 2-е, испр. и доп. - Новосибирск: Новосибирск: Сиб. унив. изд-во, 2013. - 478 с. - 2500 экз. - ISBN 5-94087-077-5
9. Инге-Вечтомов С.Г. Генетика с основами селекции. - М.: Высш. шк., 2019. - 592 с.
10. Клаг Уильям С., Каммингс Майкл Р. Основы генетики. - М.: Техносфера, 2017. - 896 с.
11. Льюин Б. Гены: Пер. с англ. - М.: Мир, 2017. - 544 с.
12. Пухальский В.А. Введение в генетику. - М.: КолосС, 2017. - 224 с. (Учебники и учеб. пособия для студентов высш. учеб. заведений)