Научноисследовательская работа студентки 4 курса Направления подготовки 06. 03. 01 Биология Профиль Биохимия
 Скачать 1.18 Mb. Скачать 1.18 Mb.
|

Определение начальных условий для распределения кислорода в гемоглобинеСогласно обзору данных, мембрана эритроцита вопреки известным предположениям не является причиной замедления диффузии кислорода. Измерение коэффициента диффузии кислорода через мембрану эритроцитов дало цифру 0.7-КГ см1 І с, что примерно в 5 раз меньше чем для воды. Чтобы объяснить фактическое замедление диффузии, коэффициент должен быть примерно в 100 раз меньше, чем у воды, т. е. порядка 1.2-10"7 елі2 /с. Как отмечает Мелдон, распространено мнение, что диффузию кислорода в эритроцит замедляет неподвижный слой молекул плазмы, прилегающий к мембране эритроцита снаружи и достигающий толщины 5 мкм. В работе [10], показано, что слой жидкости между стенкой легочного капилляра и эритроцитом не превышает в среднем 0.15 мкм. Кроме того, при прохождении через капилляр эритроцит постоянно вращается в силу чисто физических причин. Поэтому слой жидкости как препятствие диффузии кислорода можно рассматривать лишь для экспериментальных условий в искусственной среде. Препятствием диффузии может служить сама «толща» гемоглобина. Кроме того, по мнению [2] важное значение имеет различие кислородсвязывающих свойств гемоглобина в эритроците и растворе. Высокая концентрация гемоглобина в эритроците (около 33%), наличие солей, ДФГ и других веществ уменьшают его сродство к кислороду и замедляют соответствующую реакцию. В растворе все вышеперечисленные факторы не действуют. Модельные кривые хорошо описывают экспериментальными данными. В данном случае значение параметра а для процесса оксигенации составило: а = 0.9 10э м% /(с моль). Здесь также наблюдается хорошее сравнение с экспериментом. Символы -экспериментальные данные для различного начального заполнения. Кривые моделирование. В данном случае значение параметра а для процесса диссоциации составило: а = 1.3 103 м3 /(с моль). ГЛАВА 2 МЕХАНИЗМЫ РЕГУЛЯЦИИ СВЯЗЫВАНИЯ ГЕМОГЛОБИНА С КИСЛОРОДОМНаш организм потребляет в сутки примерно 400–600 л кислорода [1]; такое же его количество содержится в 100 тоннах воды. Если бы наше сердце вместо крови перекачивало жидкость с таким же содержанием кислорода, то за каждое сокращение нужно было бы прокачивать 15 л этой жидкости. Представляете себе такой насос? Реальный сердечный выброс в 100 раз меньше. Такая фантастическая экономия возникает благодаря тому, что в нашей крови может раствориться в 100 раз больше кислорода, чем в воде. Для этой цели в организме имеются специальные клетки – эритроциты. В этом обзоре мы расскажем, как устроен эритроцит, обеспечивающий необыкновенно высокую эффективность транспорта кислорода. Гемоглобин Эритроциты занимают 40% объема нашей крови. Эти клетки в норме представляют собой двояковогнутые диски, на 98% заполненные красным белком– гемоглобином, связывающим кислород (рис. 1) [2]. Гемоглобин – относительно небольшой интенсивно окрашенный белок с молекулярной массой 66 000– 68 000 Дальтон [2–5]. Гемоглобина в крови так много, что он определяет ее цвет. Молекула белка состоит из четырех похожих субъединиц, каждая из которых способна обратимо связывать кислород [2, 3]. Для этого связывания не требуется энергия. Таким образом, каждая молекула гемоглобина способна связать от одной до 4 молекул кислорода.  Рисунок 8 – Электронная микрофотография эритроцитов [2] Связывание кислорода с гемоглобином происходит в активном центре – геме (рис. 9). Гем – это изящная химическая конструкция, состоящая из четырех пятичленных гетероциклов, лежащих в одной плоскости. Четыре атома азота в этой плоской конструкции образуют в центре зону, которая хорошо связывает атом железа – главный элемент гемоглобина, который в свою очередь непосредственно связывает кислород. Химические связи железа с атомами азота не ковалентные, а координационные – более слабые. Это нужно для того, чтобы железо могло связывать кислород обратимо [2]. Электроны железа не должны перейти на кислород, иначе произойдет окислительно-восстановительная реакция между железом и кислородом. В результате мы потеряем кислород: он превратится в один из сильнейших окислителей на Земле – супероксид-радикал, способный натворить много бед. Координационные связи в силу своей слабости достаточно хорошо удерживают кислород в геме, но не отдают ему свои электроны. Еще одна важная особенность гема состоит в том, что одинарные и двойные связи в нем чередуются. В результате возникает система делокализованных электронов. Множество электронов, образующих 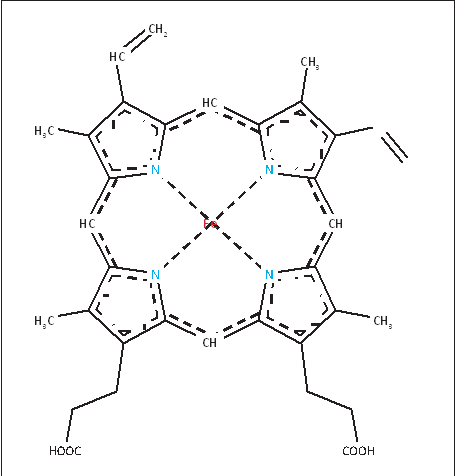 Рисунок 9 – Структура молекулы гема; пунктиром показаны связи, образующие делокализованную систему электронов [2] химические связи между атомами гема, оказываются принадлежащими всему кольцу – это создает уникальную возможность легко менять электронные свойства всей молекулы в целом и в первую очередь атома железа за счет слабых влияний соседних химических групп [2]. Возможность управлять электронным состоянием железа позволяет изменять способность железа связываться с кислородом. Атом железа может образовать 6 координационных связей; в геме четыре из них лежат в плоскости кольца и связывают железо с азотами; две другие перпендикулярны плоскости кольца (рис. 3). Одна из этих связей используется для связывания кислорода (рис. 3 Б), а противоположная ей – связь железа с азотом гистидина, занимающего 93-е положение в последовательности аминокислотных остатков глобина (так называется белковая часть молекулы гемоглобина), – как раз и позволяет управлять связыванием железа-скислородом [2, 4]. Посколькугистидин 93 является частью глобина, конформационные изменения этой молекулы могут изменять положение гистидина относительно плоскости кольца гема. В отсутствие кислорода конформация глобина такова, что гистидин 93 несколько вытягивает железо из плоскости кольца на себя (рис. 3 А). В этом состоянии гем хуже связывает кислород, чем в ситуации, когда железо находится в плоскости кольца. Таким образом, конформация белковой молекулы влияет на силу связывания кислорода. 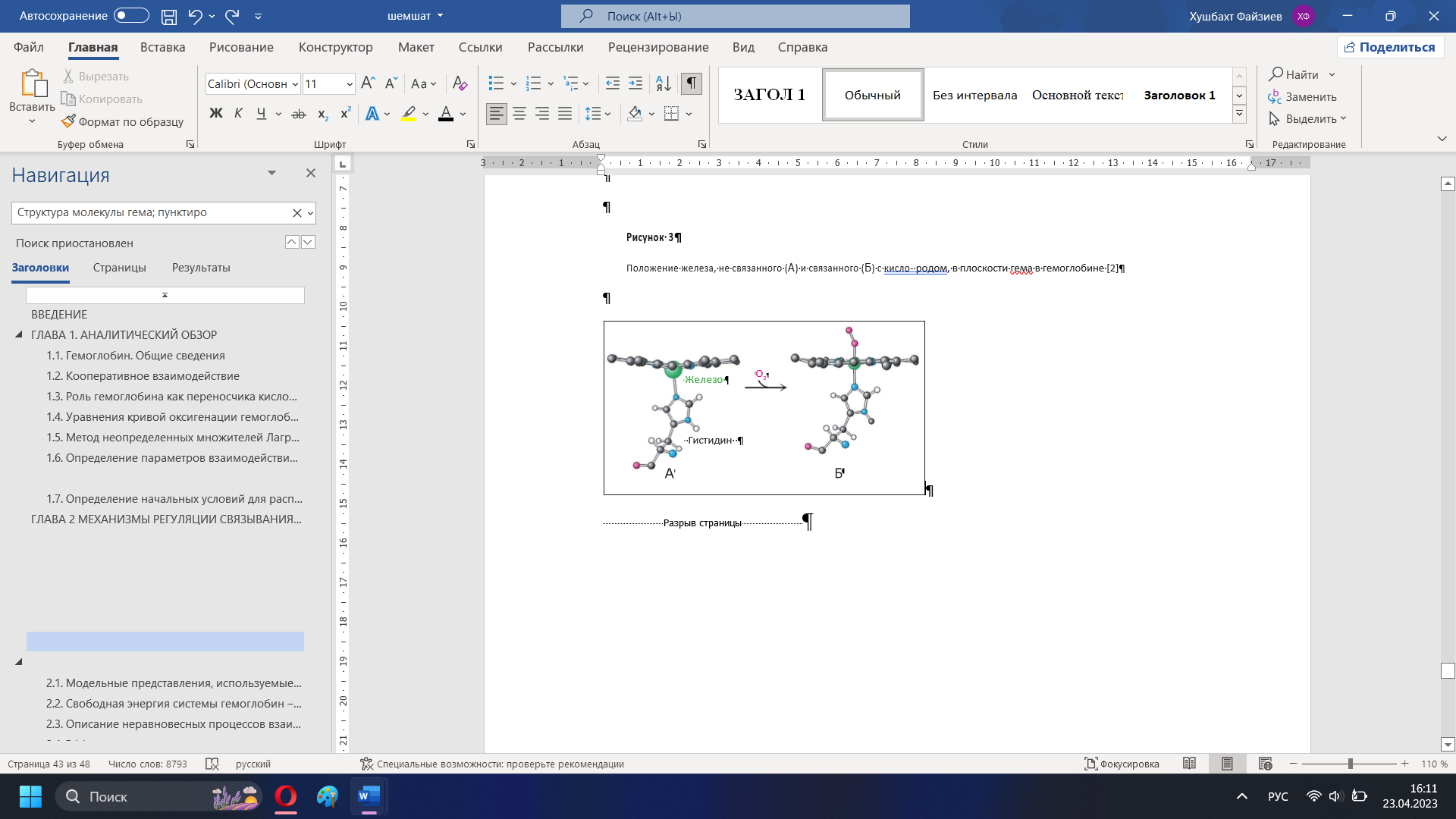 Рисунок 3 – Положение железа, не связанного (А) и связанного (Б) с кислородом, в плоскости гема в гемоглобине [2] Эта сила важна, так как организм может использовать только свободный, не связанный с гемоглобином кислород. Поэтому важно не только то, сколько кислорода есть в крови в целом и как гемоглобин его связывает, но и как он кислород отдает. Поскольку в легких растворимого кислорода больше всего, там доминирует связывание. По мере того, как кровь из легких поступает в другие органы и по капиллярам расходится по разным тканям, растворенный в крови кислород потребляется клетками, уходит из крови внутрь клеток, концентрация его уменьшается. Равновесие гемоглобина с кислородом сдвигается в сторону отдачи кислорода. Кислород освобождается и выходит в кровь [1, 6]. Молекулярная регуляция, определяющая конформацию гемоглобина и его способность связывать и отдавать кислород, позволяет более эффективно регулировать процесс переноса кислорода из легких в ткани. Рассмотрим, как это происходит (рис. 4). Кислород связывается с гемоглобином обратимо, поэтому доля гемоглобина, связанного с кислородом, зависит от давления кислорода в крови (его концентрации). Это давление в легких (100 мм рт. ст., или 100 torr) гораздо выше, чем аналогичное давление в тканях в покое (20–30 torr) [1, 2]. Организму нужен такой гемоглобин, который будет максимально связывать кислород в легких и отдавать большую его часть в тканях, ибо та часть гемоглобина, которая связала кислород и не отдала его в тканях, представляет собой балласт – мы зря гоняем груженые вагоны из легких в ткани и обратно. Закономерности этого обмена определяются двумя основными параметрами кривой связывания гемоглобина с кислородом. Эта кривая имеет S-образную форму и описана уравнением Хилла (1) [4, 7]: HbО2/Hb0 = [P]n/([P]n+[P50]n) (2), где HbO2 – концентрация гемоглобина, связанного с кислородом; Hb0 – полная концентрация гемоглобина; Р – давление кислорода в крови (величина, пропорциональная концентрации кислорода); Р50 – давление кислорода, при котором гемоглобин насыщен кислородом наполовину; n – коэффициент в формуле Хилла (1). 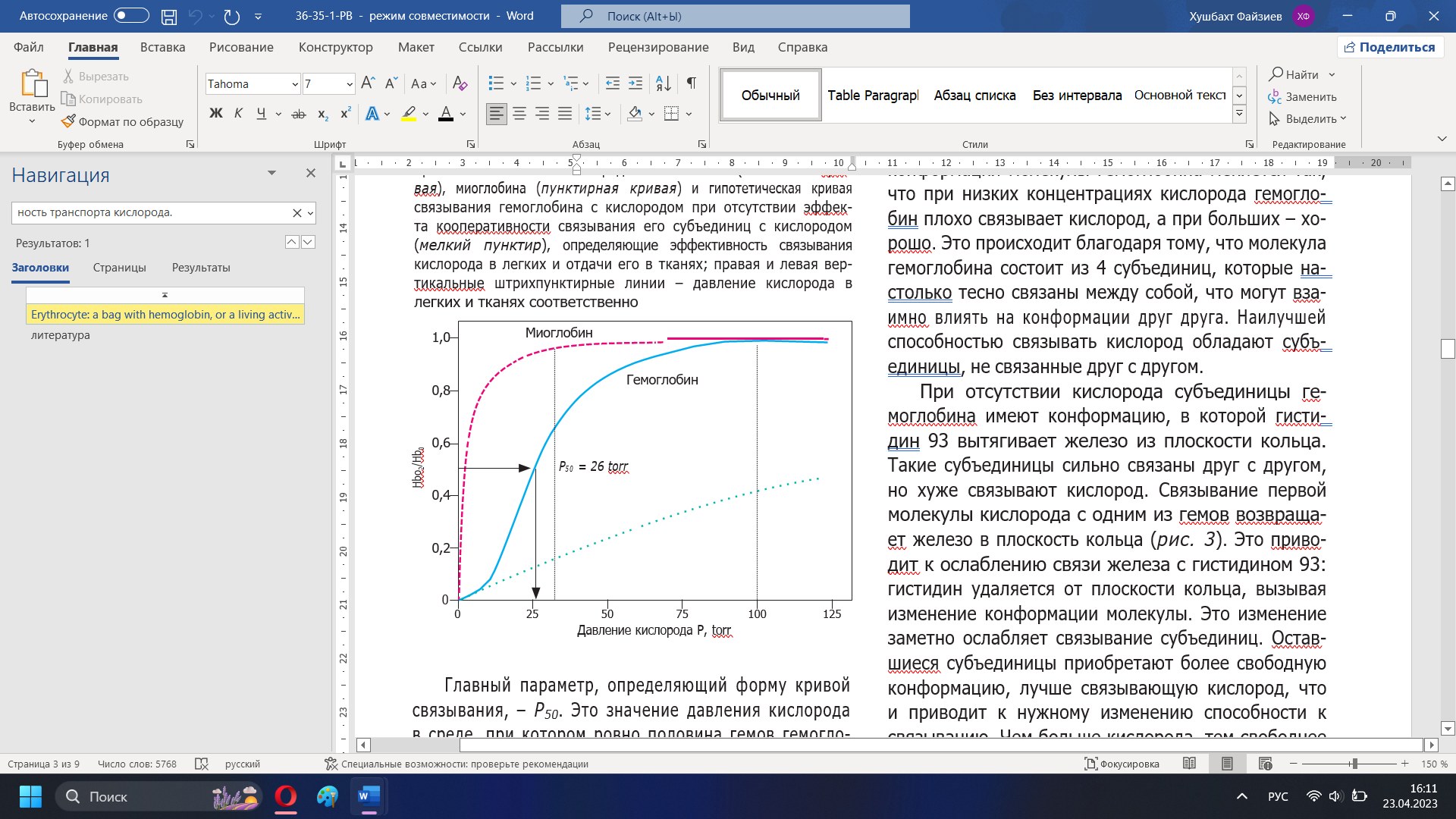 Рисунок 11 – Кривые связывания с кислородом гемоглобина (сплошная кривая), миоглобина (пунктирная кривая) и гипотетическая кривая связывания гемоглобина с кислородом при отсутствии эффекта кооперативности связывания его субъединиц с кислородом (мелкий пунктир), определяющие эффективность связывания кислорода в легких и отдачи его в тканях; правая и левая вертикальные штрихпунктирные линии – давление кислорода в легких и тканях соответственно Главный параметр, определяющий форму кривой связывания, – Р50. Это значение давления кислорода в среде, при котором ровно половина гемов гемоглобина насыщена кислородом. На рисунке 9 показано, как определяется эта величина. Второй важный параметр этой кривой – коэффициент Хилла, или коэффициент кооперативности. Как уже было сказано, кривая связывания гемоглобина с кислородом имеет S-образный характер (сплошная линия на рис. 4) и не похожа на кривую обычного химического связывания (пунктир на рис. 4). Связывание, характерное для гемоглобина, называют кооперативным. При низких концентрациях кислорода кривая с увеличением его концентрации растет медленно – гемоглобин плохо связывает кислород. Постепенно, при увеличении концентраций кислорода, кривая становится все круче. В области концентраций кислорода, близких к Р50, гемоглобин значительно эффективнее (резче) связывает или отдает кислород по сравнению с некооперативным сорбентом. S-образность характеристики помогает гемоглобину сильнее связывать кислород в легких и больше отдавать его в тканях, чем это могло бы быть при отсутствии кооперативности. Если бы S-образность была более выраженной, эффективность переноса была бы еще больше. Неполная отдача кислорода, видимо, физиологически обоснована – это запас для быстрого увеличения его потребления. Увеличение физиологической нагрузки, сопряженное с увеличенным потреблением кислорода, снизит концентрацию кислорода в тканях, что позволит отнять у гемоглобина больше кислорода. На молекулярном уровне S-образность кривой связывания гемоглобина с кислородом означает, что конформация молекулы гемоглобина меняется так, что при низких концентрациях кислорода гемоглобин плохо связывает кислород, а при больших – хорошо. Это происходит благодаря тому, что молекула гемоглобина состоит из 4 субъединиц, которые настолько тесно связаны между собой, что могут взаимно влиять на конформации друг друга. Наилучшей способностью связывать кислород обладают субъединицы, не связанные друг с другом. При отсутствии кислорода субъединицы гемоглобина имеют конформацию, в которой гистидин 93 вытягивает железо из плоскости кольца. Такие субъединицы сильно связаны друг с другом, но хуже связывают кислород. Связывание первой молекулы кислорода с одним из гемов возвращает железо в плоскость кольца (рис. 3). Это приводит к ослаблению связи железа с гистидином 93: гистидин удаляется от плоскости кольца, вызывая изменение конформации молекулы. Это изменение заметно ослабляет связывание субъединиц. Оставшиеся субъединицы приобретают более свободную конформацию, лучше связывающую кислород, что и приводит к нужному изменению способности к связыванию. Чем больше кислорода, тем свободнее субъединицы и тем лучше связывание [4]. Таким образом, субъединицы гемоглобина кооперируют в связывании кислорода – это и называют эффектом кооперативности [8, 9]. S-образность кривой связывания теряется при отсутствии эффекта кооперативности (на рис. 4 крупным пунктиром показана кривая связывания некооперативного белка миоглобина с кислородом). Миоглобин – мышечный белок, близкий родственник гемоглобина, который по структуре и способности связывать кислород очень похож на одну субъединицу гемоглобина [10]. Похожую зависимость имеет аналогичная кривая для мономеров гемоглобина. Гипотетическая кривая связывания гемоглобина с кислородом, рассчитанная при условии отсутствия эффекта кооперативности, показана мелким пунктиром на рисунке 4. Если можно было бы зафиксировать гемоглобин в этом состоянии, то кривая связывания с кислородом проходила бы очень полого. Гемоглобин умеет связывать не только кислород, но и многие другие молекулы, похожие на кислород. C этим cвязаны некоторые медицинские эффекты радикального нарушения газотранспорта. Так, место кислорода может занимать молекула оксида углерода (СО), в быту она известна как угарный газ. Эта молекула очень похожа на кислород. Она связывается с гемоглобином сильнее кислорода и очень плохо диссоциирует из образовавшегося комплекса, освобождая гемоглобин [4]. Таким образом, если во вдыхаемом нами газе появляется угарный газ, то происходит отравление: СО вытесняет кислород из молекулы гемоглобина, который оказывается загружен прочно связанным балластом и не может выполнять свою функцию, несмотря на то, что он функционально совершенно нормален. Наступает удушье. Человек, надышавшийся угарным газом, задыхается, несмотря на то, что во вдыхаемом им воздухе достаточно много кислорода. Нужно срочно избавить организм от СО – вынести отравленного человека на свежий воздух, интенсивное дыхание выводит угарный газ из кровотока. Сходный эффект вызывает другая молекула – анион цианистой кислоты (CN‾). Самая знаменитая соль этой кислоты – цианистый калий. Механизм отравления цианистым калием тот же, что и угарным газом, – блокирование связи гемоглобина с кислородом. |