Навчальнометодичний посібник з дисципліни Біологічна хімія
 Скачать 1.95 Mb. Скачать 1.95 Mb.
|

|
Номенклатура та класифікація гормонів. Гормони класифікують за місцем їх синтезу (гормони гіпоталамусу, гіпофізу, епіфізу, щитоподібної та прищитоподібних залоз, загруднинної залози, підшлункової залози, надниркових і статевих залоз), хімічною будовою, біологічними функціями та механізмом дії. За хімічною будовою (табл. 1) гормони поділяють на групи: 1. Білки: гормони передньої частки гіпофіза (крім АКТГ), інсулін, паратгормон. 2. Пептиди: АКТГ, кальцитонін, глюкагон, вазопресин, окситоцин, фактори гіпоталамуса (ліберини й статини). 3. Похідні амінокислот (адреналін, норадреналін, тироксин, трийодтиронін, гормони епіфіза). 4. Стероїдні (похідні холестерину): гормони кори надниркових залоз, статеві гормони. 5. Похідні поліненасиченої (арахідонової) кислоти - простагландини. Умовно сюди можна віднести ейкозаноїди, оскільки ці нерозчинні у воді сполуки чинять свою дію на клітини, розташовані поблизу їх місця синтезу; вони є похідними поліненасиченої арахідонової кислоти і представлені трьома класами сполук – простагландинами, тромбоксанами та лейкотрієнами. Таблиця 1. Класифікація гормонів за хімічною природою
За біологічними функціями гормони поділяють на кілька груп (табл. 2). ця класифікація умовна, оскільки одні й ті ж гормони можуть виконувати різні функції. В табл. 3 наведена класифікація за місцем синтезу гормону. Таблиця 2. Класифікація гормонів за біологічними функціями
Таблиця 3. Класифікація гормонів за місцем синтезу
За механізмом дії гормони поділяють на дві групи. До першої належать гормони, які взаємодіють із мембранними рецепторами (пептидні гормони, похідні амінокислот, а також гормони місцевої дії – цитокіни та ейкозаноїди) і контролюють процеси швидкої адаптації організму, які потребують термінового включення певного біохімічного процесу або фізіологічної функції (глікогенолізу, ліполізу, м’язового скорочення). Друга група включає гормони, які взаємодіють з внутрішньоклітинними рецепторами (стероїдні та тиреоїдині гормони), їх біологічна дія повільніша (потребує для свого проявлення декількох годин), вони відповідають за процеси довготривалої адаптації організму. Низка структурно-функціональних утворів ЦНС є одночасно і залозами внутрішньої секреції, ендокринні залози поділяють на центральні (гіпоталамус, гіпофіз і епіфіз) і периферійні (щитоподібна, підшлункова (клітини острівців Лангерганса), плацента (тимчасова ендокринна залоза періоду вагітності), тимус (або загруднинна залоза), статеві (сім’яники і яєчники), прищитоподібні, надниркові залози. Гормони також синтезуються ендокринними клітинами, що розташовуються дифузно у різних органах і тканинах, які спеціалізовані на виконання певних функцій, їх об’єднують у так звану АПУД-систему. Наприклад, ентерохромафінні клітини кишки виділяють серотонін, який регулює її функцію; базофіли сполучної тканини виділяють гістамін, а клітини нирок – ангіотензин, який бере участь у регуляції артеріального тиску. Поширені в тканинах і рідинах організму калікреїни (калідин та брадикінін) мають виражений гіпотензивний ефект завдяки своїй судинорозширювальній дії. Механізми дії гормонів Гормони впливають на тканини вибірково, що обумовлено неоднаковою чутливістю тканин до цих тканин. Органи та клітини найбільш чутливі до дії певного гормону називають мішенню гормону (орган-мішень або клітина-мішень). Концепція тканини-мішені. Тканина-мішень – це така тканина, що в якій гормон викликає специфічну фізіологічну (біохімічну) реакцію. Загальну реакцію тканини-мішені на дію гормону визначає певний ряд факторів. Перед усім це локальна концентрація гормону поблизу тканини-мішені, що залежить від: швидкості синтезу та секреції гормону; анатомічної близькості тканини-мішені до джерела гормону; констант зв’язування гормону зі специфічним білком-переносником (якщо такий є); швидкості перетворення неактивної чи малоактивної форми гормону в активну; швидкості зникнення гормону з крові в результаті розпаду чи виведення. Власне тканинна відповідь визначається: відносною активністю (чи) ступенем зайнятості специфічних рецепторів; станом сенситизації-десенситизації клітини. Специфічність гормонів по відношенню до клітин-мішеней обумовлена наявністю специфічних рецепторів. Рецептор - це одна або група білкових молекул, яка є високоспецифічною стосовно до відповідного гормону. У структурі рецептора є дві функціональні ділянки: 1) ділянка зв’язування з гормоном; 2) ділянка трансдукції (передачі) гормонального сигналу. Яким чином білок-рецептор дізнається ту молекулу гормону, з якої він може взаємодіяти? Один з доменів білка-рецептора має в своєму складі ділянку, комплементарну певній частині сигнальної молекули. Процес зв'язування рецептора з сигнальною молекулою схожий на процес утворення фермент-субстратного комплексу і може визначається величиною константи спорідненості. Більшість рецепторів вивчені недостатньо, тому що їх виділення та очищення дуже складні, а вміст кожного виду рецепторів у клітинах дуже низький. Але відомо, що гормони взаємодіють зі своїми рецепторами фізико-хімічним шляхом. Між молекулою гормону і рецептором формуються електростатичні і гідрофобні взаємодії. За зв'язування рецептора з гормоном відбуваються конформаційні зміни білка-рецептора і комплекс сигнальної молекули з білком-рецептором активується. В активному стані він може викликати специфічні внутрішньоклітинні реакції у відповідь на прийнятий сигнал. Якщо порушений синтез або здатність білків-рецепторів зв'язуватися з сигнальними молекулами, виникають захворювання - ендокринні порушення. Є три типи таких захворювань. 1. Пов'язані з недостатністю синтезу білків-рецепторів. 2. Пов'язані зі зміною структури рецептора - генетичні дефекти. 3. Пов'язані з блокуванням білків-рецепторів антитілами. Властивості рецепторів: чітка субстратна специфічність; насиченість; спорідненість з гормоном в межах біологічних концентрації гормону; зворотність дії. Клітини, які мають рецептор до гормону - це клітини-мішені відповідного гормону. Дія гормонів проявляється через їх взаємодію з рецепторами клітин-мішеней, які можуть бути сконцентровані в одній тканині або в кількох. Клітина-мішень відрізняє відповідний гормон від великої кількості інших молекул завдяки наявності на ній рецепторів із специфічним центром зв’язування з гормоном, що забезпечує їм високий ступінь вибірковості. Клітини-мішені - це клітини, які специфічно взаємодіють з гормонами з допомогою спеціальних білків-рецепторів. Рецептори локалізуються у плазматичній мембрані клітин (для гормонів пептидної природи та адреналіну) або в їх цитоплазмі (для глюкокортикоїдів) чи ядрі (для статевих і тиреоїдних гормонів). Залежно від ступеня впливу гормону на їх біологічні властивості, розрізняють гормонозалежні та гормоночутливі клітини. Прикладом гормонозалежних структур є тканини периферійних ендокринних залоз (щитоподібної, кори надниркових залоз) відносно дії тропних гормонів гіпофізу (ТТГ та АКТГ, відповідно). Гормоночутливими є клітини органів, що реагують на дію інсуліну, який контролює в них обмін глюкози, ліпідів та амінокислот (клітини м’язів, жирової тканини, лімфоїдної системи). Ефекти, які викликають гормони на клітини-мішені. Важливо відзначити, що гормони не викликають ніяких нових метаболічних реакцій в клітині-мішені. Вони лише утворюють комплекс з білком-рецептором. В результаті передачі гормонального сигналу в клітині-мішені відбувається включення або виключення клітинних реакцій, що забезпечують клітинну відповідь. При цьому в клітині-мішені можуть спостерігатись наступні ефекти: 1) зміна швидкості біосинтезу окремих білків (в тому числі білків-ферментів); 2) зміна активності вже наявних ферментів (наприклад, в результаті фосфорилювання); 3) зміна проникності мембран в клітинах-мішенях для окремих сполукабо іонів (наприклад, для Са2+). Раніше вже відмічалось про механізми впізнавання гормонів – гормон взаємодіє з клітиною-мішенню тільки при наявності спеціального білка-рецептора. Слід додати, що зв’язування гормону з рецептором залежить від фізико-хімічних параметрів середовища – від рН, концентрації різних іонів. Особливе значення кількість молекул білка-рецептора на зовнішній мембрані або всередині клітини. Типи гормональних рецепторів. Рецептори для фізіологічно активних сполук (гормонів та інших біорегуляторів) за своєю хімічною природою в більшості випадків належать до глікопротеїнів, вільні поверхні яких представлені олігосахаридними ланцюгами (глікозильні групи). Вони відповідають за розпізнавання зовнішніх сигналів. На мембрані однієї клітини можуть знаходитися різні типи рецепторів. Зв’язування гормону з рецептором здійснюється за рахунок іонних, ван-дер-ваальсових і гідрофобних взаємодій. Усі рецептори поділяються на два класи – мембранні та внутрішньоклітинні (цитозольні та ядерні), що різняться за своєю молекулярною організацією та послідовністю біохімічних реакцій, які включаються після взаємодії фізіологічно активних сполук (ФАС) із специфічними рецепторними білками. Залежно від способу передачі гормонального сигналу в клітину виділяють три класи мембранозв’язаних рецепторів (рис. 1): 1. Рецептори, які володіють каталітичною активністю – при взаємодії ліганда з рецептором активується внутрішньоклітинна частина (домен) рецептора, що має тирозинкіназну або тирозинфосфатазну, або гуанілатциклазну активність. За цим механізмом діють СТГ, інсулін, пролактин, інтерлейкіни, ростові фактори, інтерферони α, β, γ. 2. Каналоутворюючі рецептори – приєднання ліганду до рецептора викликає відкриття іонного каналу на мембрані. Таким чином, діють нейромедіатори (ацетилхолін, гліцин, ГАМК, серотонін, гістамін, глутамат). 3. Рецептори, пов’язані з G-білками – передача сигналу від гормону відбувається за допомогою G-білка. G-білок впливає на ферменти, що утворюють вторинні посередники (месенджер). Останні передають сигнал на внутрішньоклітинні білки. 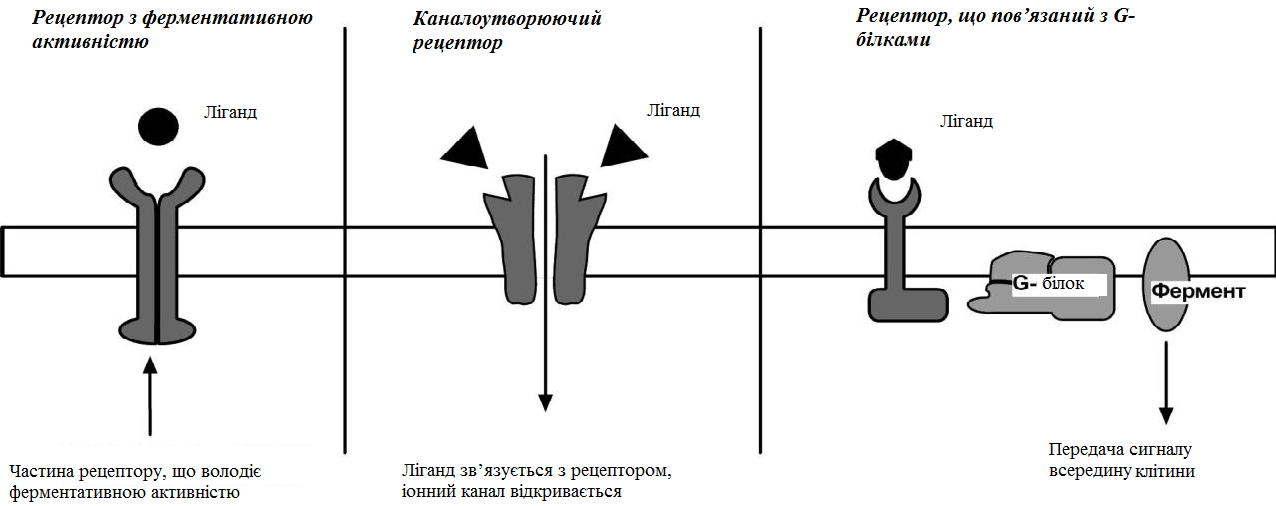 Рис. 1. Типи мембранних рецепторів До останнього класу відноситься аденілатциклазний і кальцій-фосфоліпідний механізми. 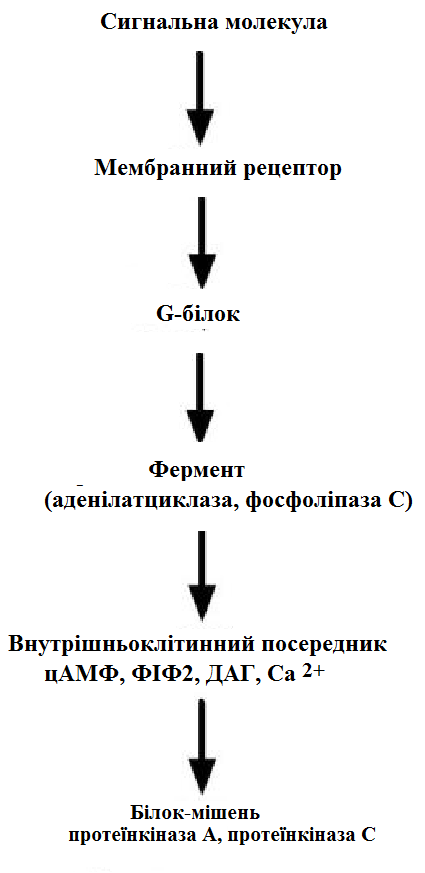 Рис. 2. Схема механізму, пов’язаного із G-білками Аденілатциклазний механізм (рис. 3) За аденілатциклазним механізмом діють гормони гіпофізу (ТТГ, ЛГ, МСГ, ФСГ. АКТГ), кальцитонін, соматостатин, глюкагон, паратгормон, адреналін (через α2 - і β-адренорецептори), вазопресин (через V2-рецептори). 1. Взаємодія ліганду з рецептором призводить до зміни конформації останнього. 2. Це зміна передається на G-білок, який складається з трьох субодиниць (α, β і γ), α-субодиниця пов'язана з ГДФ. У складі G-білків α-субодиниці бувають двох типів по відношенню до ферменту: активуючі αs і інгібуючі αi. 3. У результаті взаємодії з рецептором β- і γ-субодиниці відщеплюються, одночасно ГДФ замінюється на ГТФ. 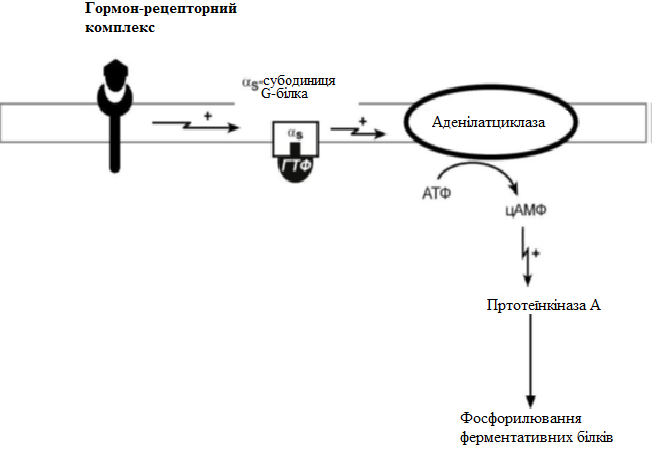 Рис. 3. Спрощена схема аденілатциклазного механізму дії гормонів 4. Активована таким чином αS-субодиниця стимулює аденілатциклазу, яка починає синтез цАМФ. 5. цАМФ – вторинний месенджер – у свою чергу, взаємодіє з протеїнкіназою А і активує її. 6. Протеїнкіназа А фосфорилює ряд ферментів, серед яких кіназа фосфорилази глікогену, глікогенсинтаза, ТАГ-ліпаза. β-Адренорецептори взаємодіють з активною субодиницею, α2-адренорецептори – з інгібуючої. Кальцій-фосфоліпідний механізм (рис. 4) За цим механізмом діють вазопресин (через V1-рецептори), адреналін (через α1-адренорецептори), ангіотензин II. Принцип роботи цього механізму (рис. 4) збігається з попереднім, але замість аденілатциклази ферментом-мішенню для α-субодиниці служить фосфоліпаза С. Фосфоліпаза С розщеплює мембранний фосфоліпід фосфатидилінозитолдифосфат до вторинних месенджерів інозитолтрифосфату (ІФ3) і диацилгліцеролу (ДАГ). Інозитолтрифосфат відкриває кальцієві канали в ендоплазматичному ретикулумі, що викликає збільшення концентрації іонів Са2+ в цитоплазмі і активацію певних кальцій-зв'язуючих білків (наприклад, кальмодуліну). Диацилгліцерол спільно з іонами Са2+ активує протеїнкіназу С, бере участь в процесах клітинної проліферації. Крім цього, диацилгліцерол має і другу сигнальну функцію: він може розпадатися на 1-моноацилглицерол і полієновую кислоту (зазвичай арахідонову), з якої утворюються ейкозаноїди. Гуанілатціклазний механізм Цей механізм поки що вивчено недостатньо докладно, але відомо, що за ним працює атріопептин (передсердний натрийуретический пептид), оксид азоту (NO) та інші, поки що мало досліджені, сполуки. Гуанілатціклазний механізм передачі сигналу в загальних рисах схожий з аденілатциклазним: після появи будь-якої сигнальної молекули сигнал сприймається ферментом гуанілатциклазою і далі передається за допомогою вторинного месенджера цГМФ. Він впливає на протеїнкіназу G, яка фосфорилює певні білки, що змінює активність клітини. Але, на відміну від аденілатциклази, даний фермент має чотири різновиди, три з яких пов'язані з мембраною, четвертий – цитозольний. Мембранозв'язані форми гуанілатциклази працюють як рецептори, що володіють каталітичною активністю. Цитозольний фермент є димером і містить у своєму складі гем, він безпосередньо взаємодіє у цитозолі з сигнальною молекулою (наприклад, оксид азоту). 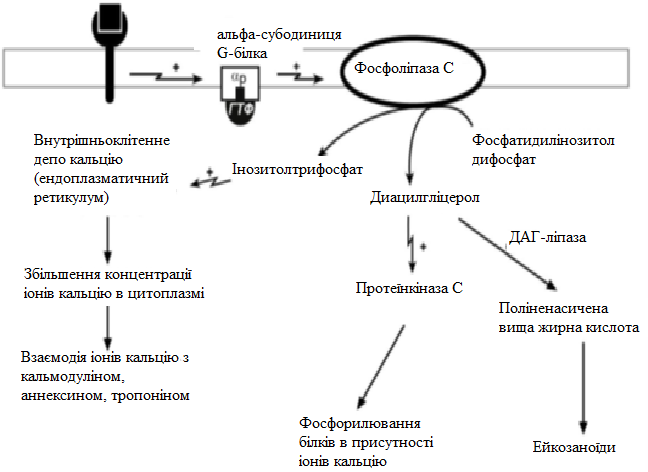 Рис. 4. Спрощена схема кальцій-фосфоліпідного механізму дії гормонів 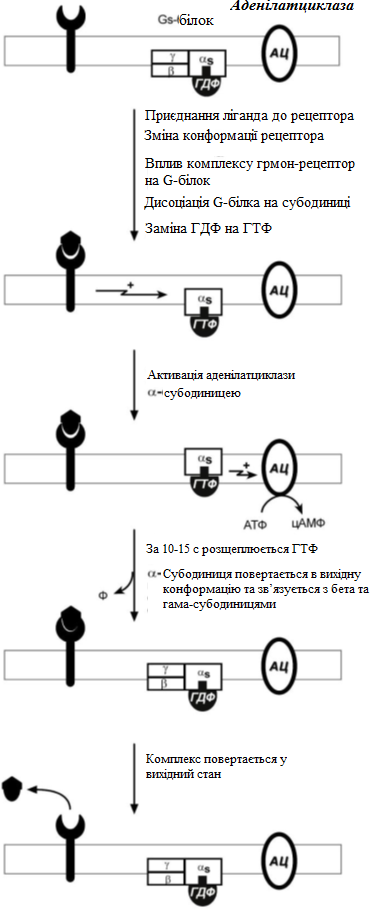 Рис. 5. Схема активації аденілатциклази при дії гормонів 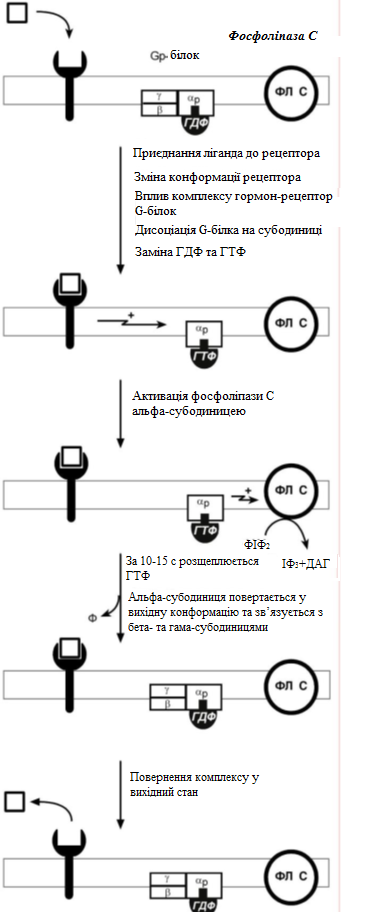 Рис. 6. Схема активації фосфоліпази С при дії гормонів Цитозольні рецептори За цитозольним механізмом діють стероїдні гормони і гормони щитовидної залози. Будучи гідрофобними молекулами, вони потрапляють в клітини шляхом простої дифузії і міцно зв'язуються з рецепторами, розташованими в цитоплазмі. Комплекс гормон-рецептор в деяких випадках додатково модифікується і активується. Далі він проникає в ядро, де може зв'язуватися з ядерним рецептором. В результаті гормон знаходить спорідненість до ДНК. Зв'язуючись з гормон-чутливим елементом в ДНК, гормон впливає на транскрипцію певних генів і змінює концентрацію РНК в клітині і, відповідно, кількість певних білків у клітці. Іонотропні рецептори – інтегральні олігомерні білки, які складаються з кількох субодиниць, між якими знаходиться заповнений молекулами води канал. За певних зовнішніх впливів (зміна потенціалу на мембрані, дія медіатора чи гормону) ці рецептори вибірково змінюють проникність мембрани для конкретного виду іонів. У результаті взаємодії з ФАС субодиниці рецептора підтримують канал у відкритому стані, чим створюють умови для транспорту іонів (Са2+, Na+, K+, Cl- тощо). За відсутності стимулу в результаті зміни орієнтації субодиниць одна відносно одної канал закривається, блокуючи рух іонів у цій ділянці. Фізіологічними лігандами для іонотропних рецепторів є нейротрансмітери (ацетилхолін, адреналін, медіаторні амінокислоти тощо), що локалізовані в синапсах нейронів і в нервово-м’язових пластинках. 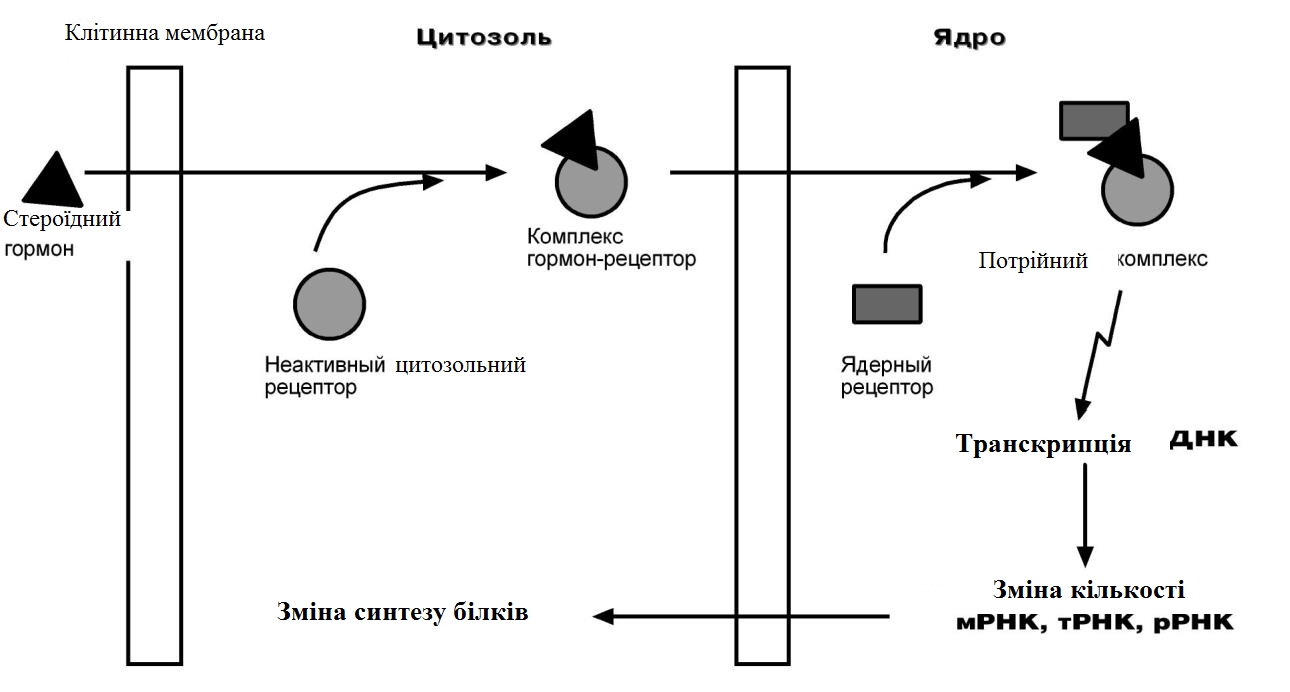 Рис. 7. Схема цитозольного механізму дії гормонів Метаботропні рецептори зв'язані не з іонними каналами, а з системами внутрішньоклітинних посередників. Зміна їх конформації після зв’язування з лігандом запускає низку біохімічних реакцій і, як наслідок, змінює функціональний стан клітини. Ці рецептори поділяють на дві групи: каталітичні рецептори, що мають власну тирозинкіназну або гуанілатциклазну активність (наприклад, рецептор інсуліну) і рецептори, що після взаємодії з ФАС викликають активацію біохімічних ефекторних систем клітини через G-білок (наприклад, рецептор вазопресину). Зв'язування ліганда (наприклад, гормона) з рецептором на зовнішній поверхні клітинної мембрани призводить до зміни активності цитоплазматичного ферменту, який, в свою чергу, ініціює клітинну відповідь (тобто інформація переноситься через мембрану). Реакція ефекторних систем клітини на дію сполук, що взаємодіють з метаботропними рецепторами, є повільною і розвивається впродовж декількох секунд. У структурі мембранних рецепторів можна виділити три функціональних ділянки: перший домен (домен впізнавання) розташовується в -кінцевій частині поліпептидного ланцюга на зовнішній поверхні клітинної мембрани, він містить глікозильовані ділянки та забезпечує впізнавання та зв’язування гормону; другий домен – трансмембранний – може складатися або з семи -спіральних поліпептидних послідовностей, або лише з одного -спірального поліпептидного ланцюга; третій (цитоплазматичний) домен створює хімічний сигнал у клітині, який забезпечує узгодженість між зв’язуванням гормону та певною внутрішньоклітинною відповіддю. Цитоплазматична ділянка рецептора до таких гормонів як інсулін, фактор росту епідермісу, інсуліноподібний фактор росту-1 на внутрішньому боці мембрани володіє тирозиназною активністю. У випадку внутрішньоклітинних рецепторів, які зв’язуються з білками-шаперонами, гормон проходить через мембрану, інформація про присутність гормону в клітині з допомогою рецептора передається в ядро. І цитозольні і ядерні рецептори стероїдних і тиреоїдних гормонів містять ДНК-зв’язуючий домен, вони теж мають три функціональні ділянки: - на С-кінці поліпептидного ланцюга рецептора знаходиться домен впізнавання та зв’язування гормону; - центральна частина рецептора включає домен зв’язування ДНК і містить амінокислотну послідовність, яка утворює 2 «цинкові пальці», у кожному з яких атом цинку зв’язаний із 4 залишками цистеїну; один «цинковий палець» відповідає за зв’язування з ДНК, а інший – за димеризацію рецепторів; - на -кінці поліпептидного ланцюга розташовується домен, який називають варіабельною ділянкою рецептора. Вона відповідає за зв’язування з тими білками, з якими бере участь у регуляції транскрипції. Деякі рецептори крім основної ділянки (або «сайту») зв’язування можуть мати додаткові регуляторні ділянки, з котрими зв’язуються інші хімічні сполуки, які змінюють реакцію рецептора на основний гормональний сигнал – посилюють або послаблюють її, або заміщають собою основний сигнал. Класичним прикладом такого рецептора є рецептор гама-аміномасляної кислоти підтипу А (ГАМК). Окрім сайту зв’язування для самої ГАМК, він має сайти зв’язування з бензодіазепінами, барбітуратами, нейростероїдами (аллопрегненолоном). мембранно-внутрішньоклітинний механізм дії гормонів для гормонів поліпептидної будови та похідних амінокислот, які не проникають усередину клітини, характерний мембранно-внутрішньоклітинний механізм дії. За фізико-хімічними властивостями це гормони, які не здатні проходити через біліпідний шар плазматичної мембрани в цитозоль. Саме тому для них рецептори знаходяться на поверхні клітини. Реалізацію дії цих гормонів у клітині забезпечують специфічні молекули, які утворюються в цитозолі у відповідь на дію гормону. Ці молекули називають месенджерами, або вторинними посередниками дії гормону. Месенджер є повноважним представником гормону в клітині. Вторинними посередниками дії гормонів можуть бути такі молекули, як: цАМФ, цГМФ, ІТФ (інозитолтрифосфат), ДАГ (диацилгліцерол), іони Са2+. Першою молекулою, що була відкрита як месенджер, став цАМФ. У 1957 р. Сазерленд відкрив, що цАМФ є вторинним посередником дії адреналіну. Нижче наведена таблиця, в якій подані гормони і відповідні вторинні посередники дії цих гормонів у клітині. Таблиця 4. |