гормоны. Гормоны-2-1. Нейроэндокринные взаимосвязи
 Скачать 484.5 Kb. Скачать 484.5 Kb.
|

|
Нейроэндокринные взаимосвязи Между эндокринной и нервной системой существует хорошо выраженная взаимосвязь, координирующим центром которой служит гипоталамус (рис. 1). Именно в нем осуществляется «переключение» с нервного пути на гуморальный. Центральная нервная система в ответ на изменение состояния внешней и внутренней среды посылает электрические сигналы в гипоталамус. В ответ на эти сигналы гипоталамус выделяет ряд гипоталамических регуляторных гормонов, которые поступают в гипофиз и регулируют секрецию его тропных гормонов. Среди гормонов гипоталамуса различают либерины, или рилизинг-факторы (рилизинг-гормоны) (от англ. release – освобождать), которые стимулируют выделение тропных гормонов гипофиза, и статины, ингибирующие этот процесс. Тропные гормоны гипофиза выделяются в кровь и осуществляют регуляцию синтеза и секреции гормонов периферическими железами. Гормоны периферических желез транспортируются с током крови и связываются с рецепторами на поверхности или внутри клеток тканей-мишеней, где они оказывают влияние на метаболические процессы. Помимо такого трансгипофизарного пути существует парагипофизарный путь, когда нервные импульсы прямо регулируют синтез и секрецию гормонов периферическими железами (например, образование адреналина в мозговом веществе надпочечников). 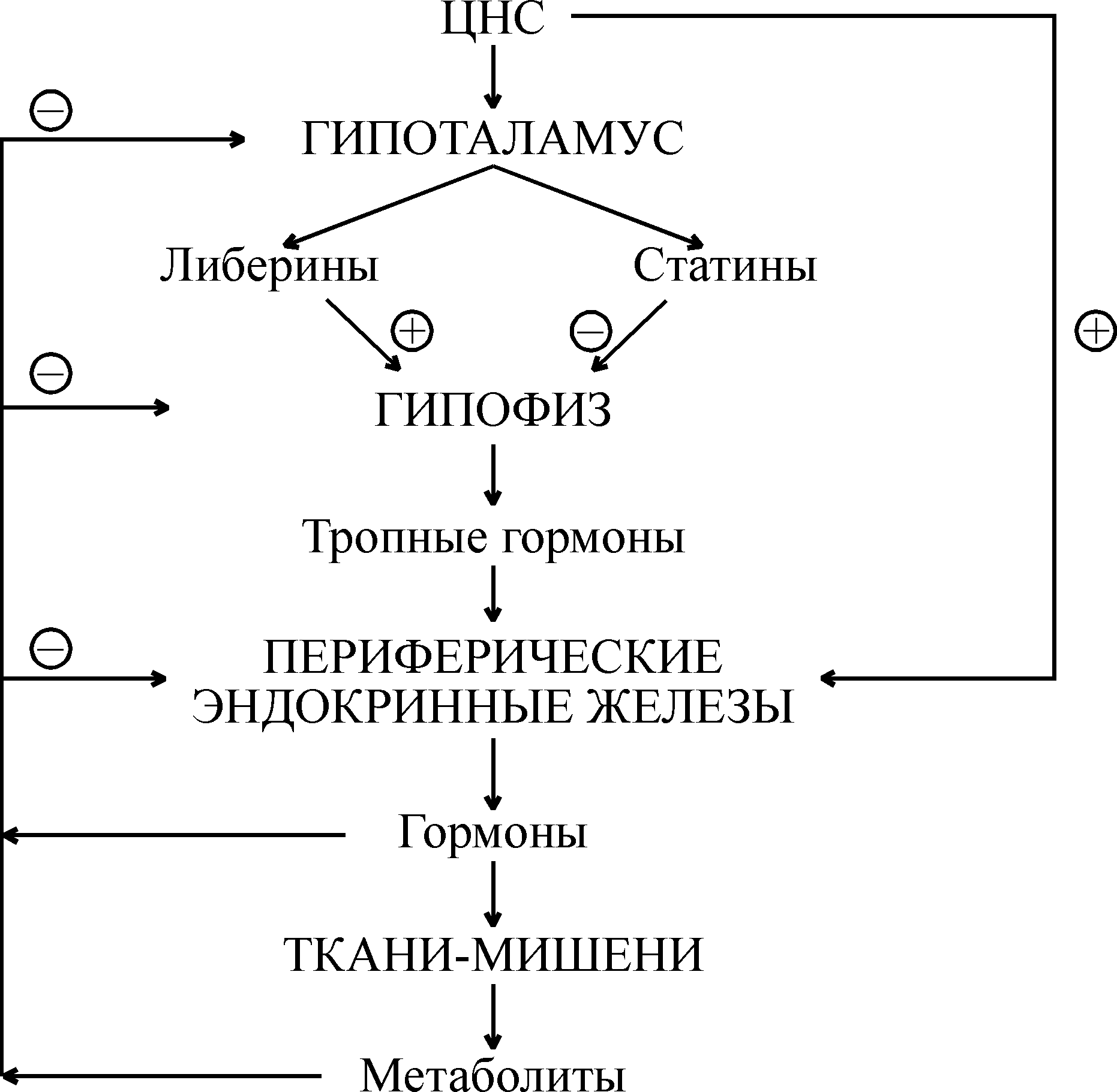 Рис. 1. Схема взаимосвязей в нейроэндокринной системе Функциональная активность эндокринной системы регулируется также с помощью механизмов, работающих по принципу отрицательной обратной связи, которая носит название «плюс-минус» взаимодействий. Гормональная обратная связь заключается в том, что при стимуляции гормонами гипофиза образования и секреции гормонов периферических желез происходит повышение уровня последних в крови (знак «плюс»), которые по механизму отрицательной обратной связи угнетают образование тропных гормонов, действуя через гипофиз или гипоталамус (знак «минус»). При снижении их концентрации происходит активация всей системы. Помимо гормональной существует также метаболитно-гормональная обратная связь: гормоны периферических желез, действуя на метаболизм в клетках тканей-мишеней, изменяют содержание тех или иных метаболитов в крови, которые оказывают влияние на секрецию гормонов прямо в периферических железах или через гипофиз и гипоталамус. Такими метаболитами могут быть аминокислоты, жирные кислоты, глюкоза, нуклеотиды и нуклеозиды, различные ионы, вода и др. Механизм действия гормонов Уровень содержания гормонов во внеклеточной жидкости очень низкий и составляет 10–7–10–12 моль/л, что намного ниже содержания других веществ. Следовательно, клетки-мишени должны отличать данный гормон от других соединений, что обусловлено наличием у этих клеток специфических рецепторов, обеспечивающих им высокую степень избирательности. Рецептор обладает очень высокой специфичностью и сродством по отношению к соответствующему гормону, которое в 1000–10000 раз выше, чем сродство к гормону других неспецифических белковых молекул. Рецепторы водорастворимых полипептидных гормонов и гормонов аминокислотной природы (за исключением тироксина), не способных проходить через клеточную мембрану, располагаются на наружной поверхности плазматической мембраны клеток тканей-мишеней. По химической природе эти рецепторы являются гликопротеинами, специфичность которых обусловлена их углеводным компонентом. Взаимодействие гормонов с рецепторами характеризуется быстрым насыщением последних, что является важным элементом механизма быстрого ответа на повышение концентрации гормона в крови. Рецепторы жирорастворимых стероидных гормонов, легко проникающих сквозь мембрану, и тиреоидных гормонов, также способных проходить через липидный бислой мембраны, локализованы в цитоплазме клеток-мишеней. Способность гормонов проникать внутрь клетки определяет молекулярные механизмы их действия. Различают следующие основные механизмы действия гормонов (и других внеклеточных регуляторов, в том числе лекарственных препаратов) на процессы обмена веществ в клетке: мембранно-внутриклеточный, мембранный и цитозольный. Мембранно-внутриклеточный механизм действия Мембранно-внутриклеточный тип действия характерен для гормонов полипептидного строения и производных аминокислот, не проникающих в клетку, в связи с чем их влияние на внутриклеточные процессы обмена опосредуется промежуточными соединениями, называемыми вторичными посредниками (первичный посредник – сам гормон). В качестве вторичных посредников могут выступать молекулы циклических нуклеотидов – циклического аденозинмонофосфата (цАМФ) и циклического гуанозинмонофосфата (цГМФ), ионы Са2+, а также продукты превращения фосфоинозитидов. Механизм действия гормонов посредством циклических нуклеотидов. Важнейшим вкладом в развитие современных представлений о вторичных посредниках было открытие цАМФ (Э.Сазерленд, 1957 г.), который впоследствии занял центральное место в схемах, освещающих механизмы действия различных биологически активных веществ – гормонов, биогенных аминов, лекарственных веществ. Обнаруженный вслед за цАМФ другой циклический нуклеотид – цГМФ – не является его простым аналогом. Пути биосинтеза цАМФ и цГМФ отличаются и реализуются через разные регуляторные системы, однако механизмы их влияния на клеточную активность сходны и сводятся к выборочному фосфорилированию функционально важных клеточных белков. Обмен и функции циклических нуклеотидов в клетках обеспечиваются комплексом ферментов, которые объединяют в аденилат- и гуанилатциклазные системы. Они включают специфические ферменты синтеза: аденилат- и гуанилатциклазы; ферменты превращения циклических нуклеотидов – цАМФ- и цГМФ-фосфодиэстеразы; цАМФ- и цГМФ-зависимые протеинкиназы и снимающие их эффект фосфопротеинфосфатазы. Аденилатциклаза катализирует образование цАМФ из АТФ, а гуанилатциклаза катализирует образование цГМФ из ГТФ. 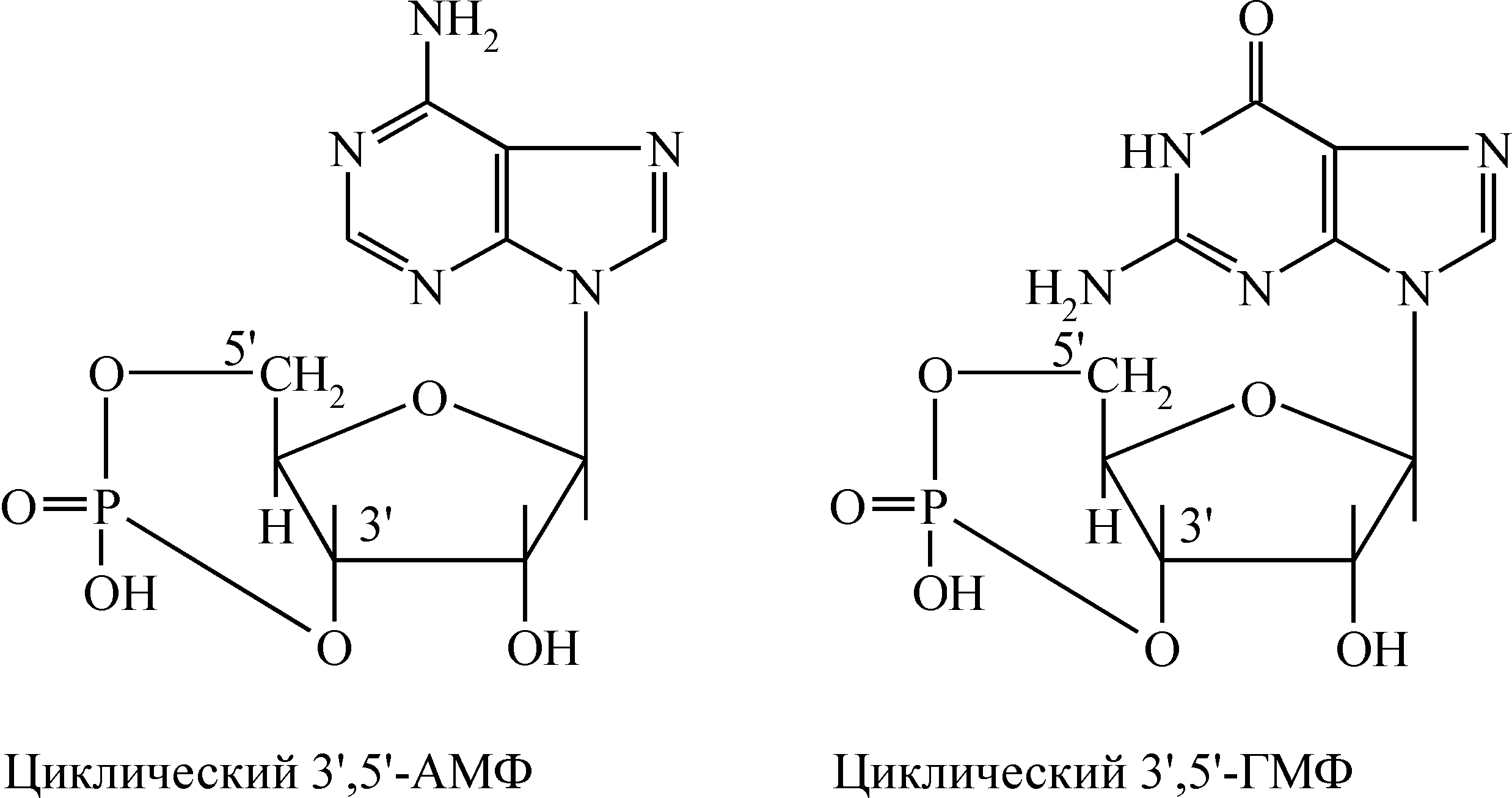 Аденилатциклаза встроена в мембрану и состоит из трех компонентов (рис. 2): первый компонент представляет собой рецептор (R), выходящий на наружную поверхность мембраны и взаимодействующий с гормоном. Второй компонент – это связующий G- или N-белок, который является ГТФ-зависимым регуляторным белком. Исследования последних 10 лет показали, что действие гормонов опосредуется не одним белком, а двумя параллельными системами – стимулирующей (s) и ингибирующей (i). Каждая система состоит из рецептора – Rs или Ri и регуляторного белка Ns (Gs) или Ni (Gi). Обе системы сопряжены с одной и той же каталитической молекулой – третьим компонентом аденилатциклазы или собственно аденилатциклазой, расположенной на внутренней поверхности мембраны и катализирующей образование цАМФ из АТФ. Взаимодействие гормона с рецептором приводит к активации либо инактивации аденилатциклазы 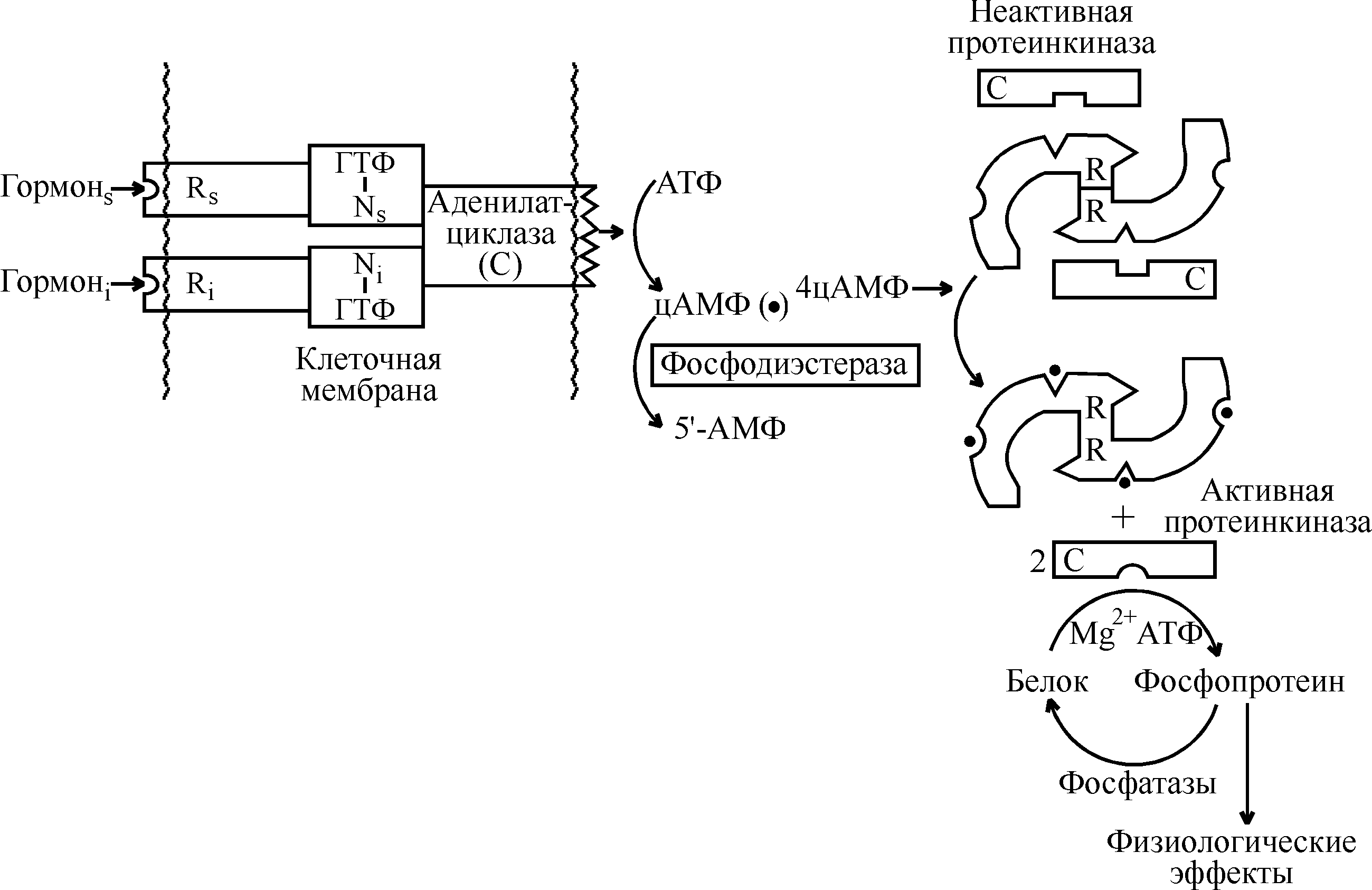 Рис. 2. Гормональная регуляция внутриклеточных процессов через цАМФ-зависимые протеинкиназы (по Марри Р., Греннер Д., Мейес П. и др.) В результате действия гормонов, активирующих аденилатциклазу, в клетке повышается уровень цАМФ. Циклический АМФ связывается с двумя регуляторными субъединицами тетрамера протеинкиназы, в результате чего происходит диссоциация регуляторных и каталитических субъединиц и тем самым активируются каталитические субъединицы, т.е. протеинкиназа переходит из неактивной формы в активную (рис. 2). Активные протеинкиназы катализируют фосфорилирование различных белков (в том числе ферментов) по схеме: Образовавшиеся фосфорилированные белки вызывают различные биологические эффекты. Функция одних белков после фосфорилирования их протеинкиназами активируется, функция других угнетается. Например, при действии адреналина и глюкагона – гормонов, увеличивающих уровень цАМФ в клетках – активированные протеинкиназы фосфорилируют гликогенфосфорилазу, переводя ее из неактивной формы в активную и обеспечивая распад гликогена в печени и скелетных мышцах. В то же время в результате фосфорилирования происходит инактивация фермента гликогенсинтетазы, что приводит к торможению синтеза гликогена. Изменение количества цАМФ в клетке под влиянием различных гормонов может осуществляться как через аденилатциклазу, так и через фосфодиэстеразу, катализирующую гидролиз цАМФ с образованием 5-АМФ. Вещества, ингибирующие фосфодиэстеразу, оказывают эффект, подобный действию гормона, а активирование фермента снижает его действие. Например, ингибиторы фосфодиэстеразы производные ксантинов – кофеин, эуфиллин, теофиллин повышают уровень цАМФ, имитируя эффект, оказываемый эндогенными гормонами. Еще один путь снятия гормонального сигнала достигается активированием фосфопротеинфосфатаз, осуществляющих дефосфорилирование белков. Больше всего накоплено сведений о роли фосфатазы в регуляции обмена гликогена в мышцах. Аналогично цАМФ гормоны и другие внеклеточные регуляторы стимулируют образование другого циклического нуклеотида – цГМФ под действием фермента гуанилатциклазы, которая существует в растворимой и мембраносвязанной формах. Активирование гуанилатциклазы приводит к образованию цГМФ из ГТФ. Циклический ГМФ активирует цГМФ-зависимые протеинкиназы, которые фосфорилируют белки. Одни белки фосфорилируются посредством цАМФ-зависимых протеинкиназ, другие – посредством цГМФ-зависимых ферментов. Поэтому в зависимости от мембранной системы, связывающей гормон и передающей сигнал в клетку, включаются цАМФ-зависимые или цГМФ-зависимые биологические процессы, имеющие часто противоположную направленность. Механизм действия гормонов посредством ионов Са2+. Ионизированный кальций является важным регулятором различных процессов, таких, как мышечное сокращение, секреция гормонов, нейромедиаторов и пищеварительных ферментов, процесс свертывания крови, возбудимость клеточных мембран. Он служит универсальным посредником, передающим внутриклеточным механизмам сигналы, поступившие в клетку извне. Клетки очень чувствительны даже к небольшим изменениям концентрации ионов Са2+. Это обусловлено тем, что их внутриклеточная концентрация очень низкая (10–7 моль/л) по сравнению с внеклеточной (10–3 моль/л). Поддержание такой разности концентраций возможно благодаря двум свойствам плазматической мембраны: низкой проницаемости для кальция и наличию специальных переносчиков – ионных насосов, которые выкачивают Са2+ из клетки против концентрационного градиента. В состоянии покоя пассивный ток кальция через плазматическую мембрану снаружи в цитозоль уравновешивается его активным транспортом в обратном направлении под действием Са2+-АТФаз или Са2+-насосов, которые за счет энергии АТФ откачивают Са2+ в обмен на Na+ или H+ из цитоплазмы во внеклеточную среду. Стимуляция клетки гормоном или нейромедиатором приводит к тому, что в плазматической мембране открываются кальциевые каналы, а также Са2+ освобождается из митохондрий и эндоплазматического ретикулума, в результате чего концентрация Са2+ в цитозоле повышается. Полагают, что быстрые эффекты кальция связаны с его мобилизацией из органелл клетки, а более поздние – с поступлением его извне или с уменьшением выхода кальция из клетки. Ионы кальция взаимодействуют с кальций-связывающим белком цитоплазмы кальмодулином. Кальмодулин содержит четыре участка связывания Са2+, и присоединение Са2+ ко всем четырем участкам приводит к значительным изменениям конформации белка, в частности, к увеличению степени спирализации и образованию компактной структуры. В результате этих конформационных переходов кальмодулин в комплексе с Са2+ приобретает способность регулировать активность определенных ферментов, что ведет к изменению метаболических процессов в клетке. Мембранный тип действия гормонов Для некоторых гормонов, не проникающих в клетку, характерен мембранный тип действия. Гормон, связываясь с рецептором на поверхности мембраны, изменяет ее проницаемость для ряда метаболитов и ионов. Это происходит в результате связывания гормона с ее транспортными системами, изменяя конформацию транспортных белков и, следовательно, проницаемость мембраны. По описанному механизму действует инсулин на мембраны жировых клеток, увеличивая их проницаемость для глюкозы. Действуя через мембранный механизм, инсулин снижает уровень глюкозы, аминокислот и некоторых ионов в крови путем увеличения проницаемости клеточных мембран. Поступление этих метаболитов в клетку оказывает влияние на биохимические процессы, а поступление ионов изменяет электрический потенциал мембран. Однако только мембранный тип действия обычно не характерен для гормонов. Наряду с локальным действием, гормоны оказывают влияние на метаболические процессы через вторичных посредников. Например, инсулин не только увеличивает проницаемость клеточных мембран, но через внутриклеточные посредники усиливает анаболические процессы в клетке. Цитозольный механизм действия Гормоны липофильной природы, молекулы которых способны проникать через липидный бислой плазматической мембраны клеток, оказывают свое действие посредством цитозольного механизма. К ним относятся все стероидные гормоны, а также иодтиронины, занимающие по липофильности промежуточное положение между стероидами и водорастворимыми гормонами. Специфические рецепторы к этим гормонам находятся в цитоплазме только клеток-мишеней. Они представляют собой белки, обладающие высоким сродством к своему гормону, благодаря стереоспецифичности связывания. Когда стероидный или тиреоидный гормон проникает через плазматическую мембрану внутрь клетки, в цитоплазме он связывается с цитозольным рецептором, образуя комплекс гормон-рецептор, который далее подвергается активации. В процессе активации изменяется конформация, величина и поверхностный заряд комплекса, и он приобретает способность проникать в ядро и связываться с определенными участками хроматина, активируя или инактивируя специфические гены. В результате влияния на транскрипцию генов изменяется содержание определенных белков, что сказывается на активности биохимических процессов в клетке. Цитозольный механизм действия называется также прямым, поскольку гормоны, способные проникать внутрь клетки, оказывают непосредственное влияние на количество ферментных белков. В отличие от цитозольного, мембранно-внутриклеточный тип действия является косвенным, так как регуляция обмена веществ, в том числе на уровне генетического аппарата, осуществляется через внутриклеточные посредники. Для стероидных и тиреоидных гормонов, оказывающих прямое влияние на генетический аппарат клетки, более характерна регуляция роста и дифференцировки тканей и органов, чем для гормонов, не проникающих в клетку. Рассмотренные выше механизмы действия гормонов – ведущие, однако в механизмах действия отдельных гормонов возникают дополнительные специфические факторы регуляции метаболизма. Гормоны периферических желез Гормоны щитовидной железы Щитовидная железа – непарный эндокринный орган, расположенный в области гортанных хрящей. Она состоит из многочисленных фолликулов, заполненных белковым коллоидом, главным компонентом которого является белок иодтиреоглобулин. Этот белок представляет собой высокомолекулярный гликопротеин (М.м. 670000), в котором содержится 0,5–1% иода и 8–10% углеводов. Он является исходным соединением, из которого образуются гормоны щитовидной железы. В щитовидной железе происходит синтез и секреция двух иодсодержащих гормонов – 3,5,3-трииодтиронина (Т3) и 3,5,3,5-тетраиодтиронина (Т4,тироксина). Кроме того, в ней образуется неиодированный гормон – кальцитонин, который вырабатывают также паращитовидные железы. Иодтиронины представляют собой иодированные производные аминокислоты тирозина: 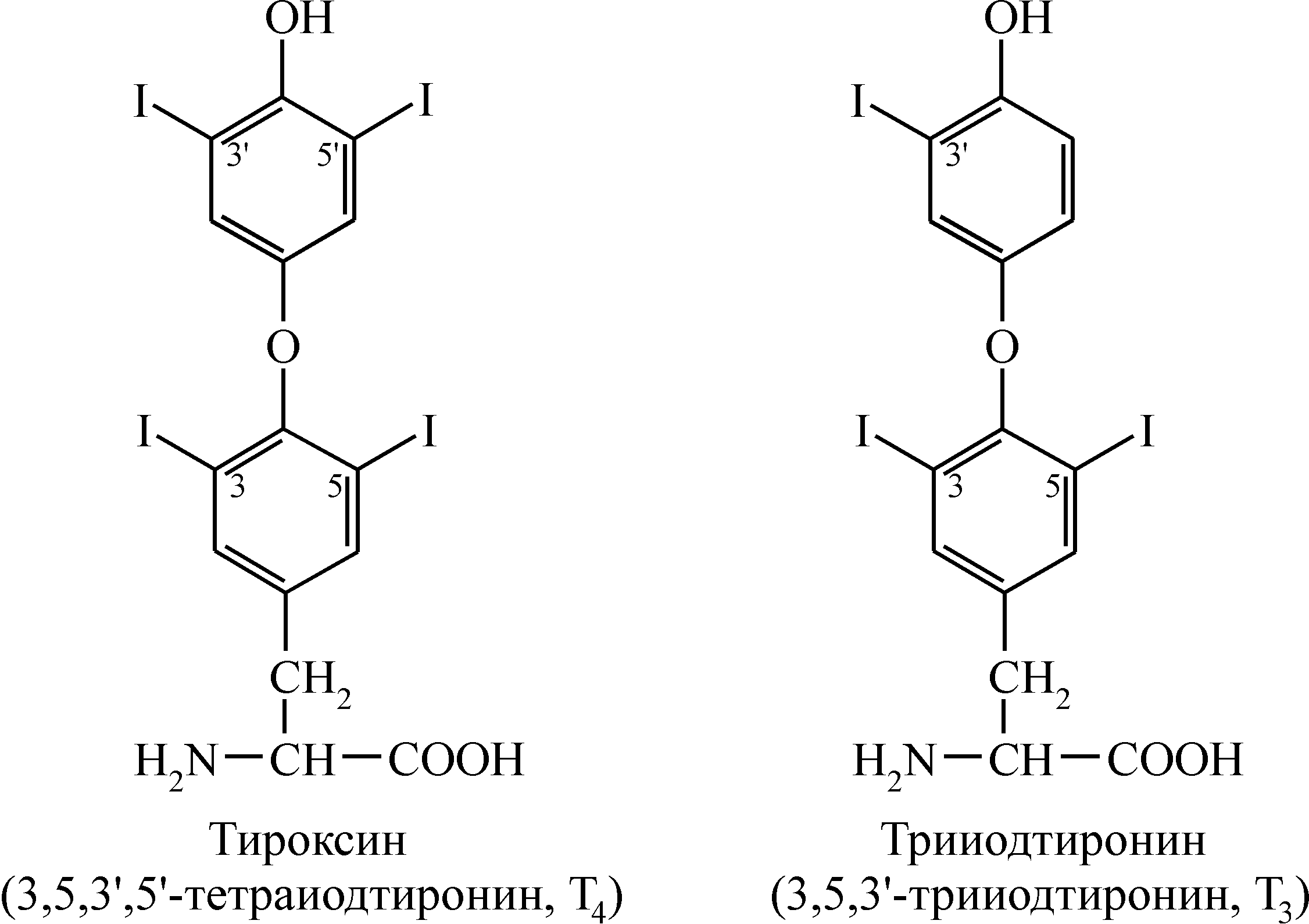 Синтез и секреция иодтиронинов Для синтеза тиреоидных гормонов необходим иодид (I–), поступающий из крови, и тиреоглобулин. Единый процесс биогенеза иодтиронинов включает следующие этапы: 1) поглощение иодидов из крови и их окисление; 2) синтез тиреоглобулина и иодирование его тирозиловых остатков; 3) образование гормональных иодтиронинов из иодированных тирозиловых остатков в молекуле тиреоглобулина; 4) протеолитическое расщепление иодтиреоглобулина и освобождение в кровь иодтиронинов. Синтез и секреция иодтиронинов регулируется гипоталамо-гипофизарной системой. Тиреолиберин гипоталамуса стимулирует секрецию тиреотропина гипофиза, который по аденилатциклазному механизму усиливает синтез и секрецию тиреоидных гормонов. Последние регулируют свой собственный синтез по механизму обратной связи. Образование тиреотропина ингибируется соматотропным гормоном гипофиза. Поступившие в кровь иодтиронины связываются с двумя белками: тироксинсвязывающим глобулином (ТСГ) и тироксинсвязывающим преальбумином (ТСПА). В количественном отношении большее значение имеет ТСГ, т.к. его сродство к гормонам в 100 раз превышает сродство ТСПА. В крови соотношение Т4 и Т3 составляет примерно 4:1, однако сродство Т3 к ТСГ значительно (в 10–100 раз) меньше, чем Т4, поэтому он быстрее поступает из крови в ткани. Этим объясняется более высокая ( в 5 раз) биологическая активность Т3 по сравнению с Т4, а также меньший период полужизни (около 2 суток) по сравнению с этой величиной для Т4 (6–7 суток). Более высокая биологическая активность Т3 объясняется также тем, что его сродство к рецепторам клеток-мишеней в 10 раз превышает сродство Т4. Поэтому Т3 является преобладающей метаболически активной молекулярной формой гормона, и в периферических тканях большая часть (80%) Т4 превращается в Т3 или реверсивный Т3, который образуется деиодированием Т4 в 5-положении. Реверсивный Т3 обладает очень слабой биологической активностью и образуется в относительно больших количествах при хронических заболеваниях. В процессе метаболизма тиреоидные гормоны в тканях подвергаются полному деиодированию, дезаминированию и конъюгации. Действие тиреоидных гормонов Тиреоидные гормоны действуют на многие ткани организма, но наиболее чувствительны к ним ткани сердца, печени, почек, скелетных мышц, в меньшей степени нервная и жировая ткани. Иодтиронины обладают широким спектром действия, в котором можно выделить два главных направления – регуляция энергетического обмена и влияние на рост и развитие организма, дифференцировку тканей. Влияние тиреоидных гормонов на энергетический обмен проявляется в повышенном поглощении кислорода большинством тканей организма, связанным с увеличением основного обмена, и продуцировании тепла, т.е. калоригенном действии. Молекулярные механизмы повышенного теплообразования пока не известны. Калоригенный эффект гормонов связывают с их влиянием на митохондрии, что выражается в увеличении размеров и количества митохондрий, а также числа крист в них. Тиреоидные гормоны повышают дыхательную способность митохондрий вследствие индукции биосинтеза митохондриальных дыхательных ферментов и активации ферментов, обеспечивающих механизм челночного транспорта водорода из цитоплазмы в митохондрии (активность митохондриальной -глицерофосфатдегидрогеназы возрастает в несколько раз). Все это увеличивает способность тканей к образованию АТФ. Гормоны щитовидной железы оказывают сложное влияние на обмен белков, углеводов и липидов, которое характеризуется двухфазным характером. Начальный эффект их действия на обмен белков проявляется в усилении синтеза белка; введение гормонов больным с недостатком тироксина понижает выведение азота из организма. При гиперфункции щитовидной железы усиливается катаболизм белков, приводящий к отрицательному азотистому балансу. Таким же двухфазным является влияние иодтиронинов на обмен гликогена: малые дозы повышают синтез гликогена, большие – усиливают его распад в печени и мышцах. Влияние тиреоидных гормонов на обмен липидов также связано с уровнем их секреции. Они усиливают синтез, мобилизацию и особенно деградацию липидов, т.е. повышают обновляемость липидов. Иодтиронины усиливают липолиз в жировой ткани и окисление жирных кислот, снижают уровень холестерина в сыворотке крови. Однако при этом стимулируется липогенез в печени за счет индукции синтеза цитоплазматической НАДФ-малатдегидрогеназы. Интенсивное окисление углеводов и липидов требует большого потребления кислорода организмом, что и наблюдается при введении гормонов. Иодтиронины регулируют также обмен витаминов и водный баланс организма, деятельность ЦНС, желудочно-кишечного тракта, функцию сердечно-сосудистой системы, восприимчивость к инфекциям. Свое действие тиреоидные гормоны осуществляют как через рецепторы, находящиеся на плазматической мембране клеток, так и через внутриклеточные рецепторы, поскольку иодтиронины могут проникать внутрь клеток. Внутриклеточные тиреоидные рецепторы обнаружены в хроматине ядра, цитозоле и митохондриях. С рецепторами главным образом связывается Т3, имеющий к ним более высокое сродство, чем Т4. Влияние тиреоидных гормонов на экспрессию генов проявляется в усилении пролиферации клеток, их роста и дифференцировки. На уровне целого организма иодтиронины являются важными модуляторами нормального роста и развития тканей и органов. Нарушения функции щитовидной железы При гипофункции щитовидной железы, или гипотиреозе, наблюдается недостаток в организме иодтиронинов. Гипотиреоз, проявляющийся с самого рождения или в раннем детском возрасте, известен под названием кретинизм. Это заболевание характеризуется выраженной физической и умственной отсталостью. Наблюдается задержка роста (такие люди имеют карликовый рост), непропорциональное телосложение, глубокие нарушения психики и крайняя умственная отсталость. При гипофункции щитовидной железы у взрослого человека развивается микседема(слизистый отек). Микседема характеризуется снижением основного обмена и температуры тела, ухудшением памяти, утолщением кожи, вследствие избыточного накопления в ней протеогликанов и воды. Гипотиреоз может наблюдаться в результате недостаточного поступления в организм иода из-за низкого его содержания в почве и воде некоторых географических регионов. Дефицит иода вызывает заболевание эндемический зоб, которое приводит к увеличению размеров щитовидной железы. Путем такого компенсаторного механизма щитовидная железа стремится поддерживать образование гормонов на нормальном уровне. Гипофункция, вызванная недостатком иодистых солей в диете, устраняется добавлением иодистого калия к поваренной соли или другим пищевым продуктам. При гипотиреозе, обусловленном нарушением биосинтеза и секреции тиреоидных гормонов, лечебный эффект достигается на основе заместительной терапии гормональными препаратами. Гиперфункция щитовидной железы, или гипертиреозхарактеризуется избыточным образованием иодтиронинов. Наиболее выраженная форма гипертиреоза получила название тиреотоксикоза, или Базедовой болезни(болезни Грейвса). Избыточная неконтролируемая продукция иодтиронинов при этом заболевании связана с повышенной скоростью синтеза иодтиреоглобулина, которая не регулируется по типу обратной связи. При тиреотоксикозе происходит усиление интенсивности основного обмена с преобладанием процессов катаболизма. Быстрое окисление жирных кислот, глицерина, углеводов требует повышенного потребления кислорода и приводит к усилению калоригенного эффекта. Характерными симптомами при тиреотоксикозе являются: увеличение основного обмена, потеря в весе, повышение температуры тела, повышение нервной возбудимости, тахикардия, пучеглазие, увеличение щитовидной железы. Гипертиреоз можно устранить хирургическим вмешательством (удалением части щитовидной железы) или лечением, которое подавляет образование гормонов. С этой целью применяются радиоактивные изотопы иода (I131) и вещества, тормозящие синтез иодтиронинов: мерказолил, перхлорат калия. Практическое применение тиреоидных гормонов В медицинской практике применяют как тиреоидин, содержащий Т3 и Т4, так и трииодтиронин, полученный синтетическим путем. Эти препараты применяются главным образом при различных формах гипотиреоза (кретинизме, микседеме), при ожирении с явлениями гипотиреоза, церебрально-гипофизарных заболеваниях, протекающих с гипотиреозом, при эндемическом зобе. |