гормоны. Гормоны-2-1. Нейроэндокринные взаимосвязи
 Скачать 484.5 Kb. Скачать 484.5 Kb.
|

|
Практическое применение кортикостероидов. В медицинской практике широко применяются синтетические препараты кортикостероидов и их производные. Получен целый ряд синтетических аналогов глюкокортикоидов (преднизолон, дексаметазон и др.), которые более активны, чем природные гормоны. Препараты глюкокортикоидов применяются при аллергических состояниях и аутоиммунных заболеваниях, таких, как ревматизм, ревматоидный артрит, коллагенозы, бронхиальная астма, нейродермиты как иммунодепрессивные, противовоспалительные, десенсибилизирующие средства. Из аналогов минералокортикоидов – наиболее широкое применение имеет дезоксикортикостерон при болезни Аддисона, миастении, общей мышечной слабости. Иммунодепрессивное действие глюкокортикоидов позволяет использовать их при трансплантации органов и тканей для подавления реакции отторжения. Однако при применении препаратов кортикостероидов может возникнуть ряд побочных эффектов, поэтому их использование в медицинской практике требует большой осторожности и четкого знания показаний и противопоказаний к применению. Гормоны половых желез Половые железы (гонады) – парные органы, представленные у мужчин семенниками, у женщин – яичниками. Семенники и яичники – железы смешанной секреции. Их экзокринная функция заключается в продуцировании половых клеток (сперматозоидов или яйцеклеток), а эндокринная – состоит в биосинтезе и секреции половых гормонов. Предшественником для синтеза всех половых гормонов является холестерин; многие стадии их образования совпадают, поэтому небольшие количества мужских и женских половых гормонов синтезируются у лиц обоего пола. Мужские половые гормоны Мужские половые железы – семенники представляют собой сложные органы, состоящие из трех основных популяций клеток: 1) клеток Лейдига, или интерстициальных клеток, синтезирующих и секретирующих в кровь мужские половые гормоны – андрогены(andros – мужчина, греч.); 2) клеток семенных канальцев, образующих сперматозоиды; 3) клеток Сертоли, создающих условия для дифференцировки и созревания половых клеток. Синтез и секреция андрогенов Главными андрогенами (С19-стероидами) являются тестостерон и дигидротестостерон (ДГТ). Биосинтез андрогенов осуществляется главным образом в семенниках и частично – в яичниках и надпочечниках. Тестостерон образуется в клетках Лейдига из холестерина и служит непосредственным предшественником половых стероидов. ДГТ образуется из тестостерона в результате восстановления кольца А под действием фермента 5a-редуктазы. Семенники вырабатывают и секретируют очень небольшое количество ДГТ, а подавляющая его часть образуется в других тканях. У мужчин суточная секреция тестостерона составляет в норме 5 мг, содержание ДГТ в плазме – 400 мкг. Транспорт андрогенов к органам-мишеням осуществляется специфическим гликопротеином плазмы, получившим название тестостерон-эстрадиол-связывающего глобулина, который обладает высоким сродством к тестостерону, ДГТ и эстрадиолу. Период полужизни тестостерона составляет около 20 мин. Действие андрогенов Ткани-мишени для андрогенов можно классифицировать в зависимости от того, подвержены ли они действию тестостерона или ДГТ. К классическим клеткам-мишеням ДГТ относятся предстательная железа, семенные пузырьки, внешние гениталии. Ткани, чувствительные к тестостерону – эмбриональный Вольфов проток, сперматогонии, мышцы, почки, кости, мозг. Тестостерон и ДГТ участвуют в процессах: 1) половой дифференцировки; 2) сперматогенеза; 3) развитии вторичных половых признаков; 4) регуляции генов и стимуляции анаболических процессов; 5) формировании психофизического статуса мужчины. Действие гормонов на органы-мишени осуществляется при помощи цитоплазматических рецепторов. Тестостерон стимулирует синтез белка в мужских половых органах. При этом повышается уровень общей клеточной РНК, включая мРНК, тРНК и рРНК. Он стимулирует репликацию ДНК в клетках-мишенях. Генерализованное анаболическое действие андрогенов на обмен белков также лежит в основе их эффектов на костную и мышечную ткани. Они усиливают общий рост организма, задерживают кальций в костях и ускоряют рост трубчатых костей в длину, формируя скелет по мужскому типу. Удлинение костей сопровождается развитием мощной скелетной мускулатуры, которое стимулируется тестостероном. Андрогены стимулируют синтез белка в почках и печени. Почки служат главной тканью-мишенью для этих стероидов, действие которых приводит к увеличению размеров почек и индукции в них синтеза ряда ферментов. Увеличение биосинтеза белков в тканях приводит к положительному азотистому балансу организма. Андрогены стимулируют развитие мужских половых органов, а в период полового созревания – формирование вторичных половых признаков – развитие хрящей гортани и формирование мужского тембра голоса, рост волос на лице и теле. Совместно с фоллитропином они активируют сперматогенез. Тестостерон оказывает существенное влияние на развитие мозга млекопитающих, участвуя в формировании нервных путей, которые контролируют поведенческие реакции и формирование психофизиологических особенностей мужчины. Такое действие гормона связано с наличием циторецепторов в различных отделах головного мозга. Индуцируя синтез белков, в том числе ферментных, андрогены вторично влияют на энергетические процессы, в частности, активируют ферменты цикла Кребса. Это повышает аэробный распад углеводов и распад липидов в тканях, увеличивая образование энергии. Гормоны повышают синтез фосфолипидов в различных клеточных мембранах и снижают содержание холестерина, но в меньшей степени, чем эстрогены. Нарушение андрогенной функции семенников Снижение уровня синтеза тестостерона называют гипогонадизмом (состояние – евнухоидизм). При гипогонадизме до полового созревания нарушается развитие вторичных половых признаков, а после полового созревания происходит их регрессия. Различают первичный и вторичный гипогонадизм. Первичный гипогонадизм обусловлен процессами, которые непосредственно влияют на семенники, в то время как вторичный – связан с нарушением секреции гонадотропинов. При гипогонадизме наблюдается недоразвитие половых органов и вторичных половых признаков, атрофия скелетной мускулатуры, чрезмерное отложение жира в подкожной клетчатке и внутренних органах, нарушение психофизиологических реакций. Практическое применение андрогенов и их аналогов. Препараты тестостерона и их синтетические аналоги применяются в клинике при гипофункции семенников и других функциональных нарушениях половой системы у мужчин. Анаболические стероиды (метиландростендиол, нероболил, ретаболил) используют при заболеваниях, протекающих с истощением, при состояниях, сопровождающихся отрицательным азотистым балансом, и при переломах для стимуляции сращивания костей. Женские половые гормоны В настоящее время известны две группы женских половых гормонов: эстрогены(от греч. oistros – страстное влечение) – С18-стероиды и прогестины– С21-стероиды. Основным эстрогеном является эстрадиол, остальные эстрогены (эстрон и эстриол) образуются при метаболизме эстрадиола. Синтез и секреция У человека главными источниками эстрогенов являются яичники и плацента, кроме того часть гормонов образуется в надпочечниках, семенниках и некоторых других тканях. Основной представитель прогестинов – прогестеронобразуется из холестерина в желтом теле и небольшое количество – в плаценте и надпочечниках. Регуляция синтеза и секреции эстрогенов и прогестерона осуществляется при участии гонадотропных гормонов гипофиза, высвобождение которых контролируется гонадолиберином гипоталамуса. Половые гормоны оказывают регуляторное действие по типу обратной связи на уровне как гипоталамуса, так и аденогипофиза. Совместное действие гонадотропных гормонов, эстрадиола и прогестерона регулирует половой цикл у женщин, который имеет три фазы: фолликулиновую, лютеиновую и инволюции желтого тела. В различные фазы полового цикла уровень секреции гормонов резко меняется. Они не накапливаются, а секретируются по мере синтеза и подобно другим стероидам связываются с транспортными белками плазмы: эстрогены транспортируются тестостерон-эстрадиол- связывающим глобулином, а прогестерон – кортикостероид-связывающим глобулином. Действие эстрогенов и прогестерона Органы-мишени эстрогенов проявляют высокое сродство к эстрадиолу, что обусловлено наличием специфических рецепторов в тканях матки, влагалища, маточных труб, молочных желез, аденогипофиза и гипоталамуса. Биологическая роль эстрогенов заключается, в первую очередь, в стимуляции роста и созревания органов размножения, а после наступления половой зрелости – в обеспечении репродуктивной функции. Гормоны оказывают влияние на развитие вторичных половых признаков в период полового созревания: развитие молочных желез, формирование скелета по женскому типу, развитие хрящей гортани и формирование характерного для женщин тембра голоса. Они влияют на формирование полового поведения и психического статуса женщины, обеспечивают процессы в фолликулиновую фазу цикла, а также протекание беременности, родов и лактации. Процесс половой дифференцировки гипоталамуса и гонадотропные функции гипофиза индуцируются эстрогенами. Действие гормонов не ограничивается органами половой сферы, они контролируют в организме функции многих органов и тканей. Основные эффекты эстрогенов, как и андрогенов, обусловлены их способностью через цитоплазматические рецепторы регулировать скорость транскрипции специфических генов. В результате взаимодействия гормон-рецепторного комплекса с негистоновыми белками хроматина стимулируется синтез новых мРНК, кодирующих специфические белки, которые оказывают влияние на метаболизм, рост и дифференцировку клеток. Эстрогены повышают скорость синтеза белка, РНК и ДНК в органах-мишенях, что приводит к увеличению размеров и числа клеток соответствующих тканей. Под влиянием эстрогенов усиливается синтез ряда специфических белков в печени: факторов свертывания крови (II, VII, IX, X), белков-переносчиков стероидных и тиреоидных гормонов, ангиотензиногена и др. Выраженное анаболическое действие эстрогенов обеспечивает положительный азотистый баланс организма. Индуцируя ферменты гликолиза и пентозофосфатного цикла, они повышают скорость окисления углеводов и образование энергии, а также облегчают процессы синтеза с участием НАДФ×Н и рибозо-5-фосфата. Влияние гормонов на липидный обмен характеризуется ускоренным обновлением фосфолипидов и снижением накопления липидов в печени и жировой ткани. Кроме того, эстрогены способствуют выведению холестерина из организма (в большей степени, чем андрогены), а также оказывают значительное влияние на метаболизм липопротеинов: повышают образование липопротеинов высокой плотности (ЛПВП) – антиатерогенных, и снижают уровень в плазме липопротеинов очень низкой плотности (ЛПОНП) – атерогенных. Возможно поэтому, у женщин в пременопаузном периоде реже развивается атеросклероз и они менее предрасположены к инфаркту миокарда, чем мужчины. Существенное влияние оказывают эстрогены на метаболизм костной ткани, повышая в ней синтез коллагена и отложение кальция и фосфора. Установлено, что тормозящее действие гормонов на Na+, K+-АТФазу мембран мышечных клеток приводит к задержке в миометрии Na+ и потере K+, в результате чего возникает деполяризация мембран миометрия, повышающая его возбудимость и сократимость. Прогестерон, подобно эстрогенам и андрогенам, проявляет первичное действие на уровне транскрипции генов. Эстрогены и прогестерон как бы дополняют регуляторные эффекты друг друга на обмен веществ, рост и развитие тканей и органов. Прогестерон действует только в период функционирования желтого тела. Его эффекты возможны на фоне предварительного или одновременного действия на ткани эстрогенов. Изменения в матке, вызванные прогестероном, облегчают фиксацию оплодотворенной яйцеклетки и имплантацию ее в слизистую матки. При наступлении беременности и развитии плода гормон тормозит сокращение матки и маточных труб, стимулирует развитие ткани молочной железы и обеспечивает лактацию. Следовательно, его действие направлено на сохранение беременности. Желтое тело вырабатывает также гормон полипептидной природы, сходный по строению с инсулином – релаксин. Эффект гормона проявляется в расширении и размягчении шейки матки, расслаблении гладкой мускулатуры самой матки и расхождении лонного сочленения (релаксации), что облегчает прохождение плода по родовому каналу. Нарушения гормональной функции яичников Состояние, вызванное снижением уровня синтеза и секреции женских половых гормонов, называется гипогонадизмом. Оно может быть обусловлено либо процессами, которые непосредственно поражают яичники и приводят к их недостаточности (первичный гипогонадизм), либо снижением гонадотропной функции гипофиза (вторичный гипогонадизм). При дефиците эстрогенов до периода полового созревания происходит задержка развития органов половой сферы, формирования вторичных половых признаков и нарушения течения половых циклов. Также наблюдаются изменения в метаболических процессах: снижается уровень кальция и фосфата, имеет место гиперлипемия, отрицательный азотистый баланс. Дефицит прогестерона приводит к нарушению половых циклов и к выкидышам. Некоторые нарушения связаны с изменением синтеза андрогенов. При синдроме поликистозных яичников (синдроме Штейна-Левенталя) гиперпродукция андрогенов приводит к гирсутизму, нарушению половых циклов и понижению фертильности. Практическое применение женских половых гормонов. В медицинской практике широкое применение получили как природные гормоны, так и синтетические препараты, обладающие гормональной активностью, но в отличие от первых, не разрушающиеся в желудочно-кишечном тракте. Эстрогены и их синтетические аналоги, а также прогестерон применяются при недостаточности яичников, нарушении половых циклов; прогестерон, кроме того, применяют для сохранения беременности. Гормоны тимуса Тимус, вилочковая, или зобная железа – непарный лимфоидно-железистый орган, расположенный у млекопитающих загрудинно, в верхнем отделе переднего средостения. Железа имеет довольно крупные размеры в детском возрасте, но к моменту полового созревания подвергается значительной инволюции. Тимус можно назвать железой смешанной секреции, так как в нем образуются лимфоидные клетки, которые «экспортируются» в периферические ткани, главным образом в лимфатические узлы и селезенку, а также гормоны, регулирующие развитие и созревание лимфоидных клеток. Из тимуса экстрагировано несколько гормонов пептидной природы. Тимозин стимулирует иммунокомпетентность Т-лимфоцитов и их пролиферацию, тимопоэтины I и II – усиливают общую дифференцировку тимоцитов; тимусный гуморальный фактор активирует ответные реакции Т-клеток на антигены, гомеостатический тимусный гормон является синергистом гормона роста и антагонистом АКТГ и тиреолиберина. Из тимуса выделено также стероидоподобное соединение тимостерин, который оказывает множественные регуляторные эффекты на лимфоидную ткань. Таким образом, тимус необходим для формирования и деятельности иммунной системы организма. При некоторых заболеваниях нормальный иммунологический статус либо нарушается, либо вообще отсутствует. Причиной этому может быть отсутствие тимуса при рождении, когда отсутствует как клеточный, так и гуморальный иммунитет (врожденная комбинированная иммунная недостаточность) или недостаточное развитие тимуса у детей. В этом случае либо нарушается синтез гуморальных антител при нормальном клеточном иммунитете (агаммаглобулинемия), либо отсутствует клеточный иммунитет при нормальном синтезе антител (синдром Ди Георга). Гормоны центральных желез Гормоны эпифиза Эпифиз, или шишковидная железа – небольшое образование, расположенное у млекопитающих между полушариями мозга. Главный гормон, вырабатываемый эпифизом, мелатонин. 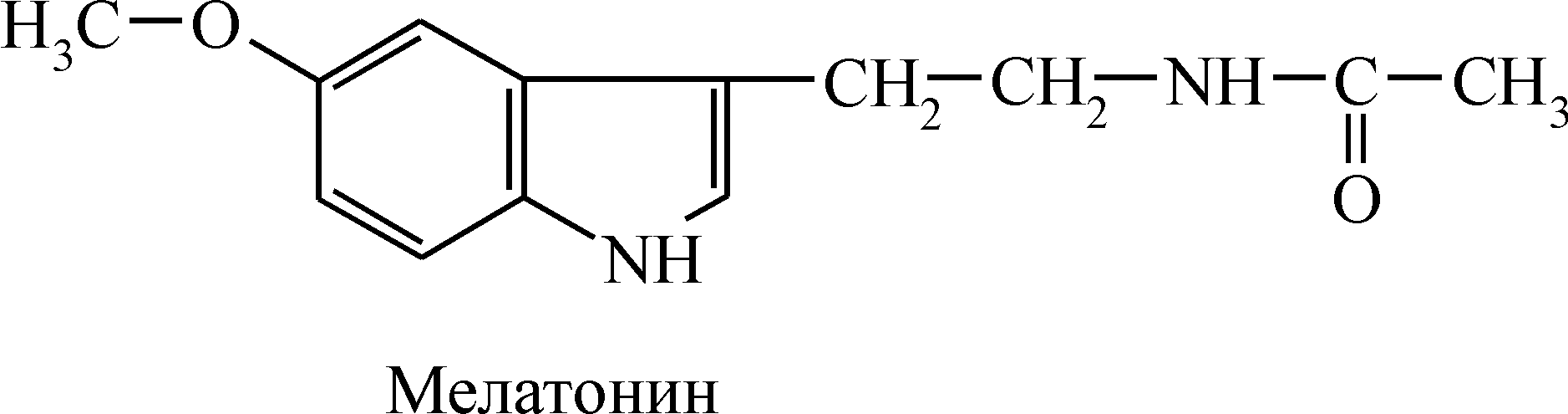 Источником для синтеза мелатонина служит аминокислота триптофан, Поэтому синтез мелатонина увеличивается ночью и снижается в дневное время. Непосредственным стимулом к повышению скорости синтеза и секреции мелатонина служит норадреналин. Основной эффект мелатонина – торможение секреции гонадотропинов, в результате чего происходит задержка полового развития. Это осуществляется либо непосредственно на уровне гипофиза, либо через гонадолиберин гипоталамуса. Увеличение светового дня тормозит синтез мелатонина и увеличивает секрецию гонадотропинов, вызывающих рост гонад и образование половых гормонов. Уменьшение светового дня снижает синтез половых гормонов и половую активность. Так как цикл биохимических процессов в железе отражает смену периодов дня и ночи, а также сезонные колебания (продолжительность светового дня зимой и летом), циклическая активность эпифиза представляет собой своеобразные «биологические часы» организма. Кроме торможения секреции гонадотропинов, мелатонин может снижать синтез и других гормонов, например, гормонов надпочечников, иодтиронинов и гормона роста. Биологическое значение этих явлений остается неизвестным. Гормон также оказывает влияние на пигментные клетки, противоположное меланотропину, – гормону средней доли гипофиза. Кроме того, мелатонин оказывает гипогликемическое, гипохолестеринемическое, антиоксидантное, противоопухолевое действие. На протяжении последнего десятилетия установлена важная роль мелатонина в процессе нейрогуморальной регуляции продолжительности жизни: торможение синтеза гормона приводит к снижению продолжительности жизни, а высокий уровень мелатонина в организме коррелирует с ее увеличением. Имеются данные об образовании в шишковидной железе еще двух гормонов: адреногломерулотропина и ингибитора гонадотропина. Нарушения гормональной функции эпифиза могут наблюдаться при опухолях. Опухоли эпифиза могут снижать половую функцию либо за счет секреции мелатонина, либо за счет разрушения нервных клеток или приводящих путей, обеспечивающих секрецию гонадолиберина гипоталамуса. С другой стороны, эти и другие близко расположенные опухоли могут повышать половую функцию, разрушая клетки эпифиза. Гормоны гипоталамуса Гипоталамус – подбугровая область промежуточного мозга, расположенная книзу от таламуса, в которой осуществляется взаимодействие высших отделов центральной нервной системы и эндокринной системы. Гипоталамус играет ведущую роль в регуляции активности аденогипофиза, а через него – в деятельности периферических желез внутренней секреции. Из гормонов, образуемых нервными клетками гипоталамуса, в настоящее время известно 6 либеринов и 3 ингибирующих гормона (статина). В табл. 19 приведены сведения об идентифицированных к настоящему времени гормонах гипофиза и гипоталамуса. По химической природе все гормоны гипоталамуса являются низкомолекулярными пептидами или олигопептидами, однако точный аминокислотный состав и первичная структура выяснены только для нескольких гормонов. Тиреолиберин представляет собой трипептид, который в отличие от классических пептидов не содержит свободной N-концевой аминогруппы и С-концевой карбоксильной группы, поскольку на N-конце находится пироглутаминовая (циклическая) кислота, а С-концевая группа амидирована. Помимо гипоталамуса гормон образуется также в других отделах нервной системы и стимулирует высвобождение не только тиреотропина, но и пролактина. Тиреолиберин оказывает влияние на поведение и терморегуляцию, действуя как антагонист опиоидных пептидов, а также является антидепрессантом. Таблица 3 Гормоны гипоталамуса и гипофиза
Соматолиберин человека – пептид, состоящий из 44 аминокислотных остатков, стимулирует синтез и секрецию гормона роста гипофиза. Соматостатинобнаружен в разных отделах головного мозга, в поджелудочной железе, клетках кишечника. Его структура и биологическое действие рассматривались ранее вместе с гормонами поджелудочной железы. Гонадолиберин, представляющий собой декапептид, контролирует высвобождение двух гормонов передней доли гипофиза – лютропина и фоллитропина. Изменение его секреции под действием сигналов, исходящих из различных отделов мозга, вызывает функциональные сдвиги, обусловленные нарушением секреции гонадотропинов. Гормон обладает антидепрессивным и возбуждающим действиями. Кортиколиберинявляется пептидом, включающим 41 аминокислотный остаток. Гормон стимулирует секрецию кортикотропина. Меланолиберинпо своей химической структуре является гексапептидом, а меланостатинпредставлен или трипептидом или пентапептидом. Меланолиберин оказывает стимулирующее, а меланостатин – ингибирующее действие на синтез и секрецию меланотропина гипофиза. Меланостатин проявляет также антидепрессивный, антинаркотический и другие психотропные эффекты. Недавно открыт 56-членный нейропептид, обладающий как гонадолибериновой активностью, так и активностью гормона, подавляющего высвобождение пролактина (пролактостатина). Его называют гонадолиберин-ассоциированным пептидом (ГАП). Химическая природа и механизм действия других гормонов гипоталамуса в настоящее время интенсивно изучается. Кроме гормонов, регулирующих функцию гипофиза, в гипоталамусе синтезируются окситоцин и вазопрессин, которые транспортируются в заднюю долю гипофиза, где запасаются и откуда секретируются. Гормоны гипофиза Гипофиз (мозговой придаток) – небольшая железа, расположенная в турецком седле основной кости черепа. Он состоит из трех долей: передней, средней и задней. Передняя и средняя доли, составляющие аденогипофиз, имеют строение железы и хорошо снабжены сосудами. Задняя доля гипофиза или нейрогипофиз, состоит из нейроглии, тесно связана с областью гипоталамуса многочисленными нервными волокнами и железистыми элементами, образующими гипофизарную ножку. Гормоны аденогипофиза В аденогипофизе вырабатываются белковые и полипептидные гормоны, большинство из которых реализует свое действие через другие периферические железы, стимулируя их развитие и функции. Поэтому гормоны гипофиза называются тропными(tropos – поворот, направление, греч.). Соматотропный гормон (гормон роста, соматотропин, СТГ)– простой белок, состоящий из 191 аминокислотного остатка. Он характеризуется высокой видовой специфичностью – в клетках человека активен только собственный гормон роста человека или высших приматов. Секреция СТГ находится под контролем гипоталамических рилизинг-гормонов – соматолиберина и соматостатина. Соматотропин необходим для линейного роста организма. Он стимулирует дифференцировку и рост тканей, способствует росту скелета, увеличению массы внутренних органов и размеров тепа. Наиболее чувствительна к СТГ хрящевая ткань, особенно расположенная в эпифизарной области трубчатых костей, в которой гормон усиливает процессы пролиферации, синтез коллагена и мукополисахаридов. Это обусловливает рост костей и всего скелета в длину. Гормон роста обладает выраженным анаболическим действием – усиливает синтез ДНК, РНК и белка во многих тканях и обеспечивает положительный азотистый баланс. Недостаточныйбиосинтез или секреция СТГ особенно опасна у детей, поскольку нарушает их способность к нормальному росту и приводит к гипофизарной карликовости. В отличие от карликового роста при гипофункции щитовидной железы, обусловливающей кретинизм, гипофизарные карлики не имеют признаков деформации скелета и не страдают умственной отсталостью. При гиперсекреции СТГ у человека, вызванной опухолью передней доли гипофиза, до периода завершения роста скелета развивается гигантизм. Если избыточная секреция соматотропина происходит после завершения роста костей, то развивается акромегалия – непропорциональный рост выступающих частей лица (носа, подбородка), увеличение размеров кистей, стоп, черепа, а также увеличение внутренних органов. Тиреотропный гормон (тиреотропин, ТТГ)является гликопротеином, который состоит из двух субъединиц – ТТГ-a и ТТГ-b. Тиреотропин регулирует функцию щитовидной железы. Он способствует поглощению клетками щитовидной железы йода и стимулирует синтез и секрецию тиреоидных гормонов. Кроме того, ТТГ в щитовидной железе повышает синтез белков, нуклеиновых кислот, фосфолипидов, вызывает увеличение размеров и количества тиреоидных клеток. Действие тиреотропина осуществляется посредством связывания со специфическими рецепторами плазматических мембран клеток щитовидной железы, в результате чего увеличивается уровень цАМФ, который и обусловливает влияние ТТГ на биосинтез тиреоидных гормонов. Адренокортикотропный гормон (кортикотропин, АКТГ) представляет собой одноцепочечный полипептид, состоящий из 39 аминокислотных остатков. Кортикотропин стимулирует синтез и секрецию глюкокортикоидов (слабо влияя на секрецию минералокортикоидов), а также обладает жиромобилизующей и меланоцитстимулирующей активностью. Липотропные гормоны (липотропины, ЛПГ)– гормоны пептидной природы, которые представлены b- и g-липотропинами. b-Липотропин стимулирует липолиз и мобилизацию жирных кислот, обладает кортикотропной, меланоцитстимулирующей и гипокальциемической активностями и, кроме того, инсулиноподобным действием, выражающимся в повышении скорости утилизации глюкозы в тканях. Стимуляция освобождения жирных кислот из жировой ткани осуществляется через аденилатциклазную систему. Физиологическая роль b-липотропина невелика и перечисленные выше биологические свойства обусловлены главным образом продуктами его распада, образующимися при ограниченном протеолизе b-липотропина: g-липотропином, a-, b-, g- и d‑эндорфинами, мет-энкефалином и b-меланоцитстимулирующим гормоном. Эндорфины и энкефалины обладают необычайной способностью подобно морфину снимать болевые ощущения. Эти пептиды, получившие название опиоиды, активно связываются с теми же рецепторами в центральной нервной системе, что и морфиновые опиаты. Они проявляют более высокую (в 18–30 раз) опиоидную активность, чем морфин. Однако роль опиоидных пептидов в организме не ограничивается их болеутоляющим действием. Они воздействуют на сердечно-сосудистую систему, умственное развитие, на процессы обучения и памяти; с ними связывают состояние эйфории и отклонения психической деятельности, которые наблюдаются вследствие повышения содержания некоторых пептидов. Кроме энкефалинов и эндорфинов обнаружено большое количество других эндогенных опиоидных пептидов – a- и b-неоэндорфины, динорфины и др. Меланоцитстимулирующий гормон (меланотропин, МСГ)секретируется средней долей гипофиза и представлен тремя типами молекул – a-, b- и g-меланотропинами. Физиологическая роль меланотропинов заключается в стимуляции у млекопитающих меланогенеза, вызывая дисперсию внутриклеточных меланиновых гранул, что приводит к потемнению кожи. Усиленная пигментация кожи у человека при низком уровне глюкокортикоидов (болезни Аддисона) может быть связана с повышенной активностью МСГ в плазме. Гонадотропные гормоны (гонадотропины). У человека аденогипофиз продуцирует два гонадотропных гормона: фолликулостимулирующий гормон (фоллитропин, ФСГ) и лютеинизирующий гормон (лютропин, ЛГ). Кроме того, в плаценте образуется хорионический гонадотропин человека (ХГЧ). Гонадотропные стимулируют рост и развитие гонад и вместе с половыми гормонами контролируют половое развитие организма и процессы размножения. Гонадотропные функции гипофиза регулируются гипоталамусом, а также в значительной мере контролируются эпифизом. Высвобождение ФСГ и ЛГ стимулируется одним гипоталамическим гормоном – гонадолиберином, секреция которого регулируется половыми гормонами по механизму обратной связи. Секреция ЛГ и ФСГ сильно варьирует в разные фазы полового цикла. Фоллитропинчерез аденилатциклазный механизм стимулирует рост фолликулов в яичниках у самок, у самцов стимулирует рост семенных канальцев и семенников, играет важную роль в инициации сперматогенеза. Лютропину самок стимулирует овуляцию и образование желтого тела в яичниках, секрецию яичниками эстрадиола и прогестерона, а у самцов – секрецию семенниками тестостерона. Гормон действует на развитие интерстициальной ткани как яичников, так и семенников. Хорионический гонадотропин человека (ХГЧ)по характеру действия сходен с гормонами гипофиза. Он появляется в крови и моче в ранний период беременности, что лежит в основе многих методов диагностики этого состояния. ХГЧ дополняет действие гормонов гипофиза при стимуляции роста желтого тела в период беременности. Лактотропный гормон (ЛТГ, пролактин, ПРЛ) представляет собой белок (М.м. 23000), состоящий из одной полипептидной цепи. Секреция пролактина находится под ингибиторным контролем нейромедиатора дофамина, к которому гипофизарные клетки имеют рецепторы. Дофамин не только снижает секрецию пролактина, но и его синтез, подавляя транскрипцию пролактинового гена. В торможении секреции пролактина участвует и гонадолиберин-ассоциированный пептид (ГАП). Что касается положительной регуляции секреции пролактина, то наличие пролактолиберина некоторые авторы ставят под сомнение. Уровень пролактина возрастает на поздних сроках беременности и при лактации. Основная функция пролактина состоит в стимуляции развития молочных желез и лактации, кроме того гормон усиливает секрецию желтым телом прогестерона, влияет на углеводный и жировой обмен. Гормоны нейрогипофиза Нейрогипофиз (задняя доля гипофиза) содержит два гормона – вазопрессин и окситоцин, которые синтезируются в нейронах гипоталамуса, откуда переносятся в заднюю долю гипофиза и поступают в кровь при соответствующей стимуляции. Оба гормона представляют собой нонапептиды, отличающиеся двумя аминокислотами в 3 и 8 положениях. Вазопрессин (антидиуретический гормон, АДГ). Нервные импульсы, вызывающие секрецию вазопрессина, являются результатом действия ряда различных стимулирующих факторов. Главные физиологические стимулы – это повышение осмоляльности плазмы и кровопотеря. Вазопрессин влияет главным образом на клетки почечных канальцев и гладкомышечные клетки сосудов. Влияние гормона на почки проявляется в мощном антидиуретическом действии: АДГ стимулирует реабсорбцию воды из гипотонической мочи в дистальных почечных канальцах и собирательных протоках, что приводит к увеличению в моче концентрации Na+,Cl-, фосфата, общего азота и повышает ее плотность. Вазопрессин стимулирует сокращение гладкой мускулатуры сосудов, оказывая сильное вазопрессорное действие. В норме он контролирует водный баланс организма и осмотическое давление плазмы крови. При дефиците вазопрессина (вызванном опухолью или инфекцией) развивается несахарный диабет, который характеризуется выделением больших объемов мочи с очень низкой удельной массой (1,002–1,006). Ежесуточно может выводиться 4–5 л мочи (полиурия). При этой патологии нарушается обратный процесс всасывания воды в канальцах почек. |