хроматин. хроматин и тд. О рганизация наследственного материала у прокариот и у эукариот
 Скачать 2.09 Mb. Скачать 2.09 Mb.
|

|
О  рганизация наследственного материала у прокариот и у эукариот рганизация наследственного материала у прокариот и у эукариот 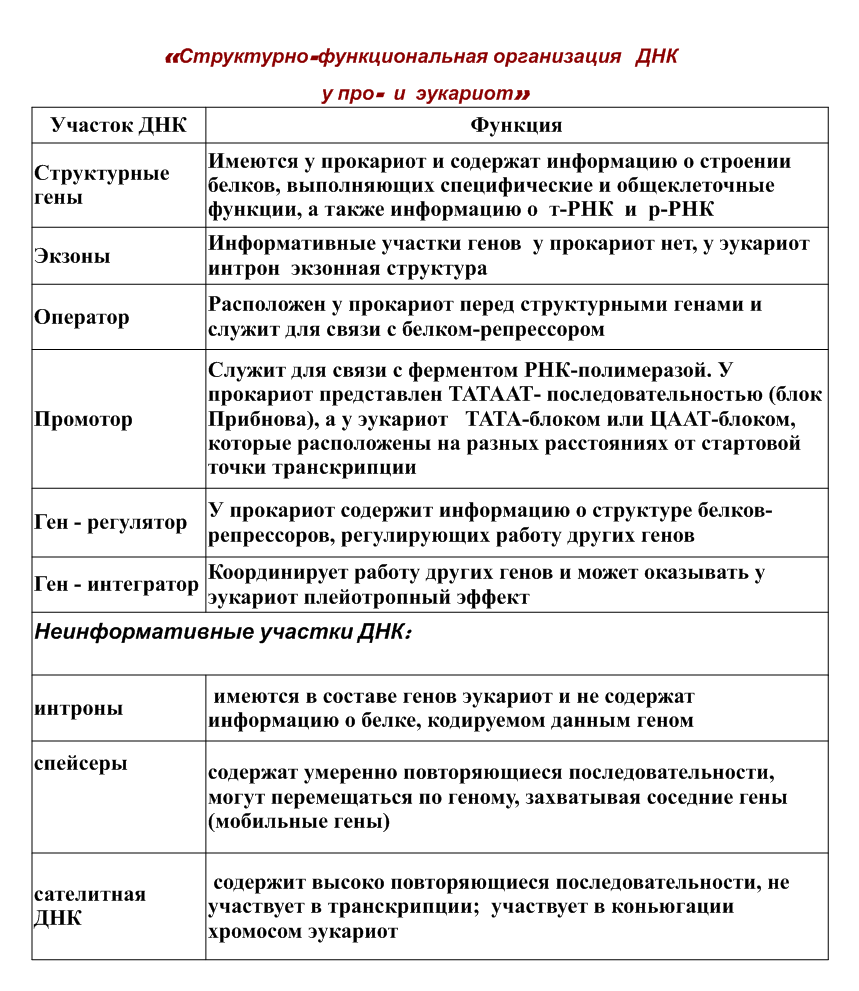   Геном современных прокариотических клеток характеризуется относительно небольшими размерами. У кишечной палочки (Е. coli) он представлен кольцевой молекулой ДНК длиной около 1 мм, которая содержит 4 • 106 пар нуклеотидов, образующих около 4000 генов. Основная масса ДНК прокариот (около 95%) активно транскрибируется в каждый данный момент времени. Как было сказано выше, геном прокариотической клетки организован в виде нуклеоида — комплекса ДНК с негистоновыми белками (см. разд. 3.5.2.4). У эукариот объем наследственного материала значительно больше. У дрожжей он составляет 2,3 • 107 п.н., у человека общая длина ДНК в диплоидном хромосомном наборе клеток — около 174 см. Его геном содержит 3 • 109 п.н. и включает, по последним данным, 30—40 тыс. генов. У некоторых амфибий и растений геном характеризуется еще большими размерами, достигающими Ю10 и 1011 п.н. В отличие от прокариот в эукариотических клетках одновременно активно транскрибируется от 1 до 10% ДНК. Состав транскрибируемых последовательностей и их количество зависят от типа клетки и стадии онтогенеза. Значительная часть нуклеотидных последовательностей у эукариот не транскрибируется вообще — молчащая ДНК. Большой объем наследственного материала эукариот объясняется существованием в нем помимо уникальных также умеренно и высоко повторяющихся последовательностей. Так, около 10% генома мыши составляют тандемно расположенные (друг за другом) короткие нуклеотидные последовательности, повторенные до 106 раз. Эти высоко повторяющиеся последовательности ДНК располагаются в основном в гетерохроматине, окружающем центромерные участки. Они не транскрибируются. Около 20% генома мыши образовано умеренными повторами, встречающимися с частотой 103—105 раз. Такие повторы распределены по всему геному и транскрибируются в РНК. К ним относятся гены, контролирующие синтез гистонов, тРНК, рРНК и некоторые другие. Остальные 70% генома мыши представлены уникальными нуклеотидными последовательностями. У растений и амфибий на долю умеренно и высоко повторяющихся последовательностей приходится до 60% генома. Избыточность генома эукариот объясняется также экзон-интрон- ной организацией большинства эукариотических генов, при которой значительная часть транскрибированной РНК удаляется в ходе следующего за синтезом процессинга и не используется для кодирования аминокислотных последовательностей белков (см. разд. 3.4.3.2). В настоящее время окончательно не выяснены функции молчащей ДНК, которая составляет значительную часть генома, реплицируется, но не транскрибируется. Высказывают предположения об определенном значении такой ДНК в обеспечении структурной организации хроматина (см. разд. 3.5.2.2). Некоторая часть нетран- скрибируемых нуклеотидных последовательностей, очевидно, участвует в регуляции экспрессии генов (см. разд. 3.6.6). Характеризуя наследственный материал прокариотической клетки в целом, необходимо отметить, что он заключен не только в нуклеоиде, но также присутствует в цитоплазме в виде небольших кольцевых фрагментов ДНК — плазмид. Плазмиды — это широко распространенные в живых клетках внехромосомные генетические элементы, способные существовать и размножаться в клетке автономно от геномной ДНК. Описаны плазмиды, которые реплицируются не автономно, а только в составе геномной ДНК, в которую они включаются в определенных участках. В этом случае их называют эписомами. В прокариотических (бактериальных) клетках обнаружены плазмиды, которые несут наследственный материал, определяющий такие свойства, как способность бактерий к конъюгации, а также их устойчивость к некоторым лекарственным веществам. В эукариотических клетках внехромосомная ДНК представлена генетическим аппаратом органелл — митохондрий и пластид, а также нуклеотидными последовательностями, не являющимися жизненно необходимыми для клетки (вирусоподобными частицами). Наследственный материал органелл находится в их матриксе в виде нескольких копий кольцевых молекул ДНК, не связанных с гистонами. В митохондриях, например, содержится от 2 до 10 копий мтДНК. Внехромосомная ДНК составляет лишь небольшую часть наследственного материала эукариотической клетки. Например, мтДНК человека содержит 16 569 п.н. и на ее долю приходится менее 1% всей клеточной ДНК. В отличие от хромосомной ДНК, мтДНК характеризуется высокой «плотностью генов». В них нет интронов, а межгенные промежутки невелики. В кольцевой мтДНК человека содержится 13 генов, кодирующих белки (3 субъединицы цитохром С-оксидазы, 6 компонентов АТФазы и др.) и 22 гена тРНК. Значительная часть белков митохондрий и пластид синтезируется в цитоплазме под контролем геномной ДНК. Если большинство ядерных генов представлены в клетках организма в двойной дозе (аллельные гены), то митохондриальные гены представлены многими тысячами копий на клетку. Для генома митохондрий характерны межиндивидуальные различия, но в клетках одного индивида, как правило, мтДНК идентична. Совокупность генов, расположенных в цитоплазматических молекулах ДНК, называют плазмоном. Он определяет особый тип наследования признаков — цитоплазматическое наследование 2. Хроматин. Виды хроматина. Хроматин является интерфазной формой существования хромосом клетки. Если хроматин упакован плотно его называют конденсированным или гетерохроматином, он хорошо видим под микроскопом. Если хроматин упакован неплотно, его называют эу- или интерхроматином. Этот вид хроматина гораздо менее плотный при наблюдении под микроскопом и обычно характеризуется наличием транскрипционной активности. Хроматин половой - хроматин, присутствующий только в женских соматических клетках и представляющий одну инактивированную Х-хромосому, которая остается в конденсированной форме. В соматических клетках женщины содержится по одной активной Х-хромосоме. По половому хроматину можно определить пол ребенка до его рождения; такое определение производится путем изучения клеток, полученных в ходе амниоцентеза, или в процессе исследования хорионических ворсинок. Существует два основных вида хроматина: 1) тельце Барра (Вагг body) - небольшое образование по краям ядра внутри ядерной оболочки, которое окрашивается при воздействии на него основных красителей; и 2) напоминающий барабанные палочки придаток ядра в нейтрофилах (разновидности белых клеток крови) Хроматин — это вещество хромосом — комплекс ДНК, РНК и белков. Хроматин находится внутри ядра клеток эукариот и входит в состав нуклеоида у прокариот. Именно в составе хроматина происходит реализация генетической информации, а также репликация и репарация ДНК. Основную массу хроматина составляют белки гистоны. Гистоны являются компонентом нуклеосом, —надмолекулярных структур, участвующих в упаковке хромосом. Классификация: 1.Эухроматин – локализующийся ближе к центру ядра, более светлый, более деспирилизованный, менее компактный, более активен в функциональном отношении. Эухроматин - неконденсированный хроматин, с которого происходит синтез белка. 2.Гетерохроматин - конденсированный хроматин, с которого белок не синтезируется. Гетерохроматин - плотно спирализованная часть хроматина, соответствует конденсированным, плотно скрученным сегментам хромосом, что делает их недоступными для транскрипции. Он интенсивно окрашивается основными красителями, и в световом микроскопе имеет вид тёмных пятен, гранул. 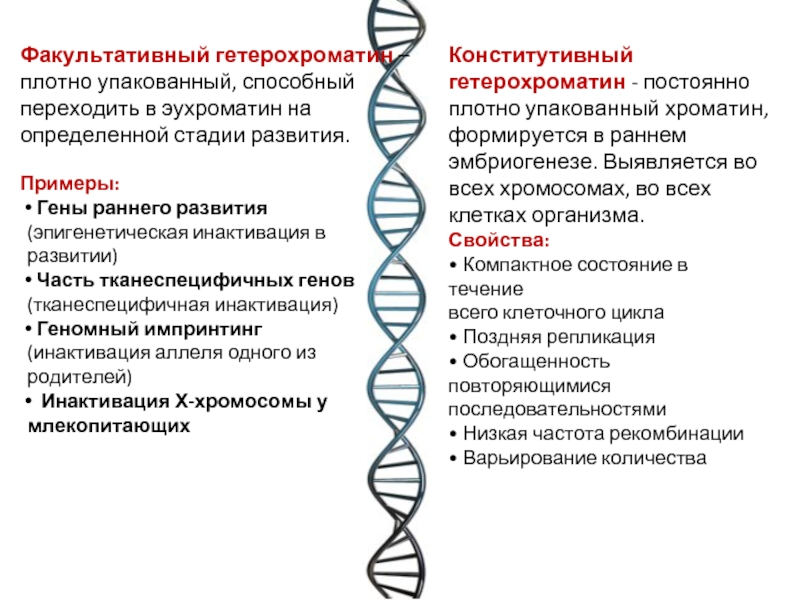 См. Учебник!!!!!!!!!!!!! 3. Строение хромосом в световом и электронных микроскопах хромосомы - наиболее упакованное состояние хроматина. В хромосомах различают первичную перетяжку (центромеру), разделяющую хромосому на два плеча. Первичная перетяжка - наименее спирализованная часть хромосомы, к ней во время деления клетки присоединяются нити веретена деления. На некоторых хромосомах есть глубокие вторичные перетяжки, отделяющие небольшие участки хромосом, называемые спутниками. В области вторичных перетяжек находятся гены, кодирующие информацию об р-РНК, поэтому вторичные перетяжки хромосом называются ядрышковыми организаторами. В зависимости от места расположения центромеры различают три типа хромосом: метацентрические (имеют плечи равной или почтиравной величины); субметацентрические (имеют плечи неравной величины); акроцентрические (имеют палочковидную форму с коротким, почти незаметным вторым плечом); Концы плеч хромосом называются теломерами Строение хромосомы наиболее хорошо удаётся рассмотреть под световым микроскопом в процессе деления клетки, а конкретно в метафазе митоза. Хромосома состоит из двух сестринских хроматид, представляющих собой нити молекулы ДНК с белками. Хроматиды образуются в результате удвоения хромосомы в процессе деления клетки. У каждой хромосомы имеется участок ДНК, называемый центромерой (кинетохором). найдено на bingoschool.ru Элементарной структурой хромосомы, различаемой с помощью электронного микроскопа, является нить, диаметром 10-13 нм, представляющая собой комплекс ДНК и гистоновых белков. Эта нить состоит из гистонового остова (в виде цепочки расположенных друг за другом белковых телец дисковидной формы), поверх которого спирально закручена нить ДНК. Комплекс ДНК и гистонов на уровне одного дисковидного тельца называется нуклеосомой. 4. Уровни компактизации наследственного материала Уровни компаюпизации хроматина: 1. Нуклеосомный - Два с половиной витка двойной спирали ДНК (в 146-200 пар нуклеотидов) наматываются снаружи на белковый кор, образуя нуклеосому. Каждый гистон представлен двумя молекулами. ДНК наматывается на кор снаружи, образуя два с половиной витка. Участок ДНК между нуклеосомами называется линкером и имеет протяжбенность 50-60 пар нуклеотидов. Толщина нуклеосомной нити составляет 8-11 нм. 2. Нуклеомерный. Нуклеосомная структура закручивается, образуя суперспираль. В её образовании принимает участие ещё один гистоновый белок HI, лежащий между нуклеосомами и связанный с линкером. К каждому линкеру присоединяется 1 молекула гистона HI. Молекулы HI в комплексе с линкерами взаимодействуют между собой и вызывают суперспирализацию нуклеосомной фибриллы. В результате образуется хроматиновая фибрилла, толщина которой составляет 30 нм (ДНК компактизирована в 40 раз). Суперспирализация происходит двумя способами. 1) нуклеосомная фибрилла может образовывать спираль второго порядка, которая имеет форму соленоида; 2) 8-10 нуклеосом образуют крупную компактную структуру - нуклеомеру. Этот уровень не допускает синтеза РНК с нуклеомерной ДНК (транскрипция не происходит). 3. Хромомерный (петельная структура). Хроматиновая фибрилла образует петли, которые сцепляются между собой с помощью особых негистоновых белков, либо петельные центры - хромомеры. Толщина 300 нм. 4. Хромонемный - образуется в результате сближения хромомеров по длине. Хромонема содержит одну гигантскую молекулу ДНК в комплексе с белками, т.е. фибриллу дезокси-рибонуклеопротеина - ДНП (400 нм) . 5. Хроматидный - хромонема складывается несколько раз, образуя тело хроматиды (700 нм). После репликации ДНК хромосома содержит 2 хроматиды. 6. Хромосомный (1400 нм). Состоит из двух хроматид. Хроматиды соединены центромерой. При делении клетки хроматиды расходятся, попадая в разные дочерние клетки. 5. Понятие о кариотипе, идиограмме, их характеристика у человека в норме. Совокупность признаков хромосомного набора, число размер и форма хромосом — кариотип. Идеограмма — это систематизированный кариотип. Хромосомы расположены по мере убывания их величины. В медицинской генетике с помощью идиограммы диагностируют некоторые хромосомные болезни. Кариотип — совокупность признаков (число, размеры, форма и т. д.) полного набора хромосом, присущая клеткам данного биологического вида (видовой кариотип), данного организма (индивидуальный кариотип) или линии (клона) клеток. Диплоидный набор хромосом, свойственным организмам данного вида. Кариограмма - это те же хромосомы метафазной пластинки, но расположенные упорядоченно. Принцип упорядоченности общий для всего вида и определяется идеограммой. Хромосомы подразделяют на аутосомы ( одинаковые у обоих полов) и гетерохромосомы, или половые хромосомы, ( разный набор у мужских и женских особей). Кариотип человека содержит 22 пары аутосом и 2половые хромосомы: ХХ у женщин и ХY у мужчин( 44+ХХ и 44+xy). В соматических клетках организмов содержится диплоидный – 2n(двойной) набор хромосом, а в гаметах – гаплоидный -1n)одинарный) Идиограмма - схематическое обобщённое изображение кариотипа с соблюдением усреднённых количеств, отношений между отд. хромосомами и их частями. На И. изображаются не только морфол. признаки хромосом, но и особенности их первичной структуры, спирализации, р-ны гетерохроматина и др. Сравнит, анализ И. используется в кариосистематике для выявления и оценки степени родства разл. групп организмов на основании сходства и различия их хромосомных наборов. Хромосомы человека принято подразделять по их размерам на 7 групп (А, В, С, D, E, F, G). Если при этом легко отличить крупные (1, 2) хромосомы от мелких (19, 20), метацентрические от акроцентрических (13), то внутри групп трудно различить одну хромосому от другой. Метафазные хромосомы человека различаются в основном по длине и расположению первичной перетяжки (центромеры). Совокупность хромосом, расположенных попарно в порядке уменьшения размера, называется кариограммой. Изображая кариограмму, половые хромосомы располагают отдельно от остальных хромосом: справа в нижнем ряду. 6. Политентные хромосомы ПОЛИТЕНИЯ (от поли … и лат. taenia - лента), наличие в ядрах некоторых соматических клеток гигантских многонитчатых (политенных) хромосом. Результат многократных удвоений хромосом, не сопровождающихся клеточным делением. Обнаружена в слюнных железах ряда двукрылых, а также у некоторых растений и простейших. Чтобы уловить изменения в структуре хроматина на уровне индивидуальных генов, необходимо изучать растянутые интерфазные хромосомы. В обычных клетках это невозможно, потому что нити интерфазного хроматина слишком тонки и запутаны. Благодаря же явлению политении на интерфазных хромосомах отчетливо видны многочисленные поперечные полосы, частота чередования которых позволяет предположить, что они соответствуют индивидуальным генам. Политенные хромосомы (гигантские хромосомы) содержат во много раз больше ДНК, чем обычные. Они не изменяют своей формы на протяжении всего митотического цикла и достигают длины до 0,5 мм, а толщины до 25 мкм. Они встречаются, например, в слюнных железах двукрылых (мух, комаров), в макронуклеусе инфузории и в тканях завязи бобов. Чаще всего они видны в гаплоидном числе, т.к. гомологичные хромосомы бывают тесно спарены. Клетки с такими хромосомами вырастают до необычно большого размера. Возникают политенные хромосомы вследствие многократно повторяющегося процесса редупликации ДНК. При этом разные участки ДНК редуплицируются в разной степени. Большинство генетически информативных областей реплицируются 1000 раз, а некоторые - более чем 30 тыс. раз. При этом циклы редупликации ДНК не сопровождаются делением клетки. По существу, политенные хромосомы представляют собой пучки множества неполностью разделенных, тесно прилежащих друг к другу индивидуальных хроматиновых нитей. В частности, политенные хромосомы слюнных желез личинки Drosophila содержат 1024 таких нити. Итак, интерфазные политенные хромосомы хорошо видны в световой микроскоп, хроматиновые петли в них располагаются в линейном порядке, при окрашивании этих хромосом заметны перемежающиеся поперечные полосы: темные - диски и светлые - междисковые участки. Предполагают, что именно диски содержат 1024 плотно уложенных гомологичных петли индивидуального петельного участка и расположенные там гены. Структурная организация и функция ДНК междисковых участков пока что неизвестна. С началом транскрипции генов диски, в которых они содержатся, декомпактизуются, становятся как бы вздувшимися и называются пуфами. Образующая их ДНК упакована гораздо менее плотно. По всей видимости, подобные структурные модификации хроматина, когда происходит его частичная деконденсация, и являются первым этапом активации эукариотических генов. Биохимически пуфы содержат меньше гистона HI, много РНК-полимеразы и, как минимум, один общий негистоновый белок. Возможно, функциональная единица генома и у высших кариот, в том числе у человека, устроена и функционирует так же. Политенные хромосомы — это гигантские хромосомы, обнаруживаемые в интерфазных клетках некоторых тканей у мух и других двукрылых. Такие хромосомы есть у них в клетках слюнных желез, мальпигиевых сосудов и средней кишки. Они содержат сотни нитей ДНК, которые редуплицировались, но не подверглись расхождению. При окраске в них выявляются четко выраженные поперечные полосы или диски (см. рис. 3.56). Многие отдельные полосы соответствуют местоположению отдельных генов. Ограниченное число определенных полос в некоторых дифференцированных клетках образует вздутия, или пуфы, выступающие за пределы хромосомы. Эти вздутые участки находятся там, где гены наиболее активны в отношении транскрипции. Было показано, что клетки разного типа содержат разные пуфы (см. рис. 3.65). Изменения в клетках, происходящие в ходе развития, коррелируют с изменениями в характере пуфов и синтезом определенного белка. Других примеров визуального наблюдения генной активности пока нет. 7. Современная теория гена   |