Обмен углеводов
 Скачать 1.99 Mb. Скачать 1.99 Mb.
|

|
Биологическое значение процесса гликолиза заключается прежде всего в образовании богатых энергией фосфатных соединений. На первых стадиях гликолиза затрачиваются 2 молекулы АТФ (гексокиназная и фосфофруктокиназная реакции). На последующих образуются 4 молекулы АТФ (фосфоглицераткиназная и пируваткиназная реакции). Таким образом, энергетическая эффективность гликолиза в анаэробных условиях составляет 2 молекулы АТФ на одну молекулу глюкозы. *КЛЮЧЕВЫЕ РЕАКЦИИ-фосфофруктокиназная, гексокиназная *преобладание изоферментов ЛДГ4 и ЛДГ5 обуславливает интенсивный анаэробный гликолиз с быстрым превращением пирувата в молочную кислоту. 3) В процессе гликолиза образуется АТФ. Суммарное уравнение гликолиза можно представить следующим образом: АТФ образуется за счёт 2 реакций субстратного фосфорилирования. Поскольку глюкоза распадается на 2 фосфотриозы, то с учётом стехиометрического коэффициента, равного 2, количество моль синтезированного АТФ равно 4. Учитывая 2 моль АТФ, использованных на первом этапе гликолиза, получаем конечный энергетический эффект процесса, равный 2 моль АТФ. Таким образом, 10 цитозольньгх ферментов, катализирующих превращение глюкозы в пируват, вместе с лактатдегидрогеназой обеспечивают в анаэробном гликолизе синтез 2 моль АТФ (на 1 моль глюкозы) без участия кислорода 4) Гексокиназа, ингибируется продуктом своей же реакции —глюкозо-6-фосфатом,а активируется субстратом — глюкозой. Регуляция гексикиназы связана с регуляцией фосфофруктокиназы: если фосфофруктокиназа ингибирована, накапливается её субстрат —фруктозо-6-фосфат,который быстро превращается вглюкозо-6-фосфат в ходе обратной реакции с помощью фосфоглюкоизомеразы. Фосфофруктокиназа является важнейшим регуляторным ферментом гликолиза. Мышечная фосфофруктокиназа аллостерически ингибируется высокими концентрациями АТФ, который связывается с регуляторным сайтом, расположенным вне активного центра, и снижает сродство фермента к субстрату —фруктозо-6-фосфату. В результате скорость реакции падает. АМФ оказывает обратное действие, повышая сродство фосфофруктокиназы к субстрату. Таким образом, скорость реакции (и гликолиза в целом) повышается. Пируваткиназа. катализирует последнюю реакцию аэробного гликолиза. Мышечный изотип этого фермента ингибируется АТФ,ацетил-КоАи аланином, а активируетсяфруктозо-1,6-бисфосфатоми фосфоенолпируватом. Печёночная пируваткиназа дополнительно ингибируется путем фосфорилирования под действием глюкагона. Биохимический смысл этой реакции в том, чтобы снизить потребление глюкозы печенью в период голодания, когда глюкоза нужна мышцам и нервным клеткам. 5) Инсулин. Печень является основным эффекторным органом панкреатических гормонов. Инсулин оказывает множественное действие на печень: он активирует гликолиз, синтез гликогена, жирных кислот и белка, ингибирует глюконеогенез, а также распад перечисленных выше соединений. Инсулин активирует транскрипцию генов глюкокиназы, фосфофруктокиназы, пируваткиназы и бифункционального фермента в печени. Кроме того, инсулин активирует ферменты фосфофруктокиназу1и пируваткиназу, ускоряя при этом весь гликолиз в целом. Глюкагон оказывает противоположное инсулину действие. Он стимулирует захват печенью аминокислот, активирует глюконеогенез, ингибирует гликолиз и биосинтез жирных кислот. Глюкагон ингибирует транскрипцию трех регулируемых ферментов гликолиза и активирует продукцию двух ферментов глюконеогенеза. Сигнальный каскад, запускаемый глюкагоном, приводит к фосфорилированию пируваткиназы и БИФ и, следовательно, к активации фруктозобисфосфатазы и ингибированию фосфофруктокиназы-2.Фруктозобисфосфатаза расщепляет фруктозо-2,6-бисфосфат,в результате чего снижается скорость гликолиза и запускается глюконеогенез . Адреналин ингибирует гликолиз в печени, подобно глюкагону. Однако он оказывает слабовыраженное активирующее действие на гликолиз в мышцах. Кроме этого, адреналин активирует распад жиров (липолиз), гликогена и глюконеогенез. Кортизол ингибирует гликолиз и активирует катаболические процессы в организме: распад гликогена, жиров, белков. 5. Охарактеризуйте процесс аэробного гликолиза по плану: 1) в каких тканях, когда и с какой целью процесс идет наиболее активно; 2) ход реакций (субстраты, ферменты / класс ферментов, продукты или характеристика основных этапов), формулы исходного субстрата и конечного продукта; 3) способы синтеза АТФ, энергетический выход в случае работы малат-аспартатного челнока; 4) аллостерическая регуляция скорости процесса (назовите аллостерические ферменты, аллостерические активаторы и ингибиторы, объясните механизм регуляции); 5) гормональная регуляция скорости процесса (назовите гормон-активатор, ключевые ферменты и возможные способы активации ключевых ферментов при участии данного гормона). 1) Гликолиз – последовательность ферментативных реакций, приводящих к превращению глюкозы в пируват с одновременным образованием АТФ. При аэробных условиях пируват проникает в митохондрии, где полностью окисляется до СО2 и Н2О. Если содержание кислорода недостаточно, как это может иметь место в активно сокращающейся мышце, пируват превращается в лактат. В тканях с аэробным метаболизмом (ткани сердца, почек и др.) преобладают изоферменты ЛДГ1 и ЛДГ2. Эти изоферменты ингибируются даже небольшими концентрациями пирувата, что препятствует образованию молочной кислоты и способствует более полному окислению пирувата (точнее, ацетил-КоА) в цикле трикарбоновых кислот. Аэробный гликолиз – процесс окисления глюкозы до пировиноградной кислоты, протекающий в присутствии кислорода. Все ферменты, катализирующие реакции этого процесса, локализованы в цитозоле клетки. 2) В аэробном гликолизе можно выделить 2 этапа: Подготовительный этап, в ходе которого глюкоза фосфорилируется и расщепляется на две молекулы фосфотриоз. Эта серия реакций протекает с использованием 2 молекул АТФ. Этап, сопряжённый с синтезом АТФ. В результате этой серии реакций фосфотриозы превращаются в пируват. Энергия, высвобождающаяся на этом этапе, используется для синтеза 10 мольАТФ. 1-фосфорилирование: 2 3 (определяет скорость гликолиза)    4-расщепление на две фосфотриозы 5-изомеризация триозофосфатов 6-окисление 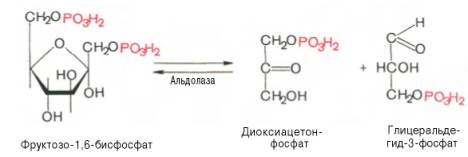  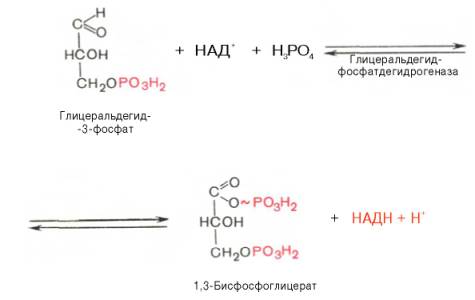 7 8-внутримолекулярный перенос фосфатной группы 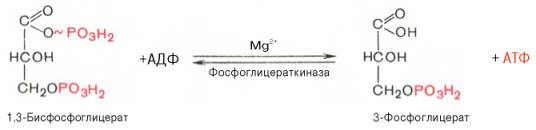  9 10-субстратное фосфорилирование 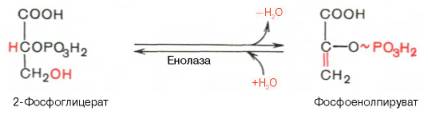  3) Реакции, связанные с синтезом АТФ, происходят после распада глюкозы на 2 молекулы фосфотриозы, т.е. на втором этапе гликолиза. На этом этапе происходят 2 реакции субстратного фосфорилирования и синтезируются 2 молекулы АТФ (реакции 7 и 10). Кроме того, одна молекула глицеральдегид-3-фосфата дегидрируется (реакция 6), а NADH передаёт водород в митохондриальную цепь переноса электронов, где синтезируется 3 молекулы АТФ путём окислительного фосфорилирования 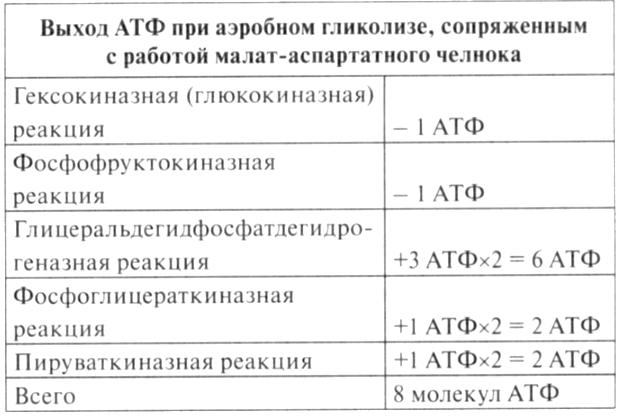 4) Гексокиназа, ингибируется продуктом своей же реакции —глюкозо-6-фосфатом,а активируется субстратом — глюкозой. Регуляция гексикиназы связана с регуляцией фосфофруктокиназы: если фосфофруктокиназа ингибирована, накапливается её субстрат —фруктозо-6-фосфат,который быстро превращается вглюкозо-6-фосфат в ходе обратной реакции с помощью фосфоглюкоизомеразы. Фосфофруктокиназа является важнейшим регуляторным ферментом гликолиза. Мышечная фосфофруктокиназа аллостерически ингибируется высокими концентрациями АТФ, который связывается с регуляторным сайтом, расположенным вне активного центра, и снижает сродство фермента к субстрату —фруктозо-6-фос-фату. В результате скорость реакции падает. АМФ оказывает обратное действие, повышая сродство фосфофруктокиназы к субстрату. Таким образом, скорость реакции (и гликолиза в целом) повышается. Пируваткиназа. катализирует последнюю реакцию аэробного гликолиза. Мышечный изотип этого фермента ингибируется АТФ,ацетил-КоАи аланином, а активируетсяфруктозо-1,6-бисфосфатоми фосфоенолпируватом. Печёночная пируваткиназа дополнительно ингибируется путем фосфорилирования под действием глюкагона. Биохимический смысл этой реакции в том, чтобы снизить потребление глюкозы печенью в период голодания, когда глюкоза нужна мышцам и нервным клеткам. 5) Инсулин. Печень является основным эффекторным органом панкреатических гормонов. Инсулин оказывает множественное действие на печень: он активирует гликолиз, синтез гликогена, жирных кислот и белка, ингибирует глюконеогенез, а также распад перечисленных выше соединений. Инсулин активирует транскрипцию генов глюкокиназы, фосфофруктокиназы, пируваткиназы и бифункционального фермента в печени. Кроме того, инсулин активирует ферменты фосфофруктокиназу1и пируваткиназу, ускоряя при этом весь гликолиз в целом. Глюкагон оказывает противоположное инсулину действие. Он стимулирует захват печенью аминокислот, активирует глюконеогенез, ингибирует гликолиз и биосинтез жирных кислот. Глюкагон ингибирует транскрипцию трех регулируемых ферментов гликолиза и активирует продукцию двух ферментов глюконеогенеза. Сигнальный каскад, запускаемый глюкагоном, приводит к фосфорилированию пируваткиназы и БИФ и, следовательно, к активации фруктозобисфосфатазы и ингибированию фосфофруктокиназы-2.Фруктозобисфосфатаза расщепляет фруктозо-2,6-бисфосфат,в результате чего снижается скорость гликолиза и запускается глюконеогенез . Адреналин ингибирует гликолиз в печени, подобно глюкагону. Однако он оказывает слабовыраженное активирующее действие на гликолиз в мышцах. Кроме этого, адреналин активирует распад жиров (липолиз), гликогена и глюконеогенез. Кортизол ингибирует гликолиз и активирует катаболические процессы в организме: распад гликогена, жиров, белков. 6. Охарактеризуйте и напишите формулами окислительную реакцию гликолиза: 1) назовите субстраты, продукты, фермент / класс фермента и его группу, кофермент-витамин; 2) охарактеризуйте дальнейшие превращения восстановленного кофермента и объясните механизм работы малат-аспартатного челнока: назовите субстрат для передачи водорода от восстановленного кофермента, фермент и продукты реакции в цитоплазме и митохондриях; 3) объясните последующее окисление восстановленного кофермента в матриксе митохондрий, приводящее к синтезу АТФ: назовите ферменты, коферменты и кофакторы цепи переноса электронов от восстановленного кофермента на конечный акцептор, фермент реакции синтеза АТФ, энергетический выход. 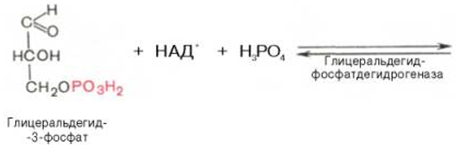 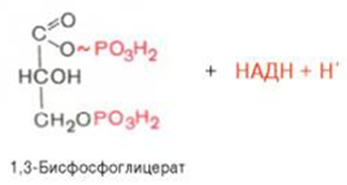 1) Субстрат – Глицеральдегид-3-фосфат Продукт – 1,3 – Бисфосфоглицерат Фермент – Глицеральдегидфосфатдегидрогеназа (класс Оксидоредуктазы, подкласс – Анаэробные дегидрогеназы) Кофермент – НАД+ Витамин – РР/B6 (Никотиновая кислота) 2) НАДН, образующийся при окислении глицералльдегид-3-фосфата, подвергается окислению путём переноса атомов водорода в митохондриальную дыхательную цепь. Т.к. цитозольный НАДН не может проникнуть в митохондрию, то перенос водорода происходит с помощью челночных систем. В малат-аспартатном челноке участвуют малат, цитозольная НАДН, митохондриальная малатдегидрогеназа. 1. В цитоплазме НАДН восстанавливает оксалоацетат в малат. (Малат с помощью переносчика попадает в митохондрию) 2. Малат окисляется в оксалоацетат НАД-зависимой малатдегидрогеназой 3. Восстановленный в ходе этой реакции НАД отдает водород в цепь переноса электронов 4. Т.к. образованный из малата оксалоацетат выйти самостоятельно из митохондрии в цитозоль не может(мембрана митохондрий для него непроницаема), то он превращается в аспартат, который выходит в цитозоль (с помощью глутамата и аспартатаминотрансферазы) 5. Аспартат превращается в оксалоацетат. Субстрат: малат Ферменты: малатдегидрогеназа – Оксидоредуктазы: Анаэробные дегидрогеназы аспартатаминотрансфераза – Трансферазы: Аминотрансферазы Продукт: цитоплазматический НАД+ 3) НАДН поступает в НАДН-дегидрогеназный комплекс (I) цепи переноса электронов. Комплекс I окисляет НАДН и восстанавливает одну молекулу убихинона, которая высвобождается в мембрану. На каждую окисленную молекулу НАДН комплекс переносит через мембрану четыре протона. НАДН-дегидрогеназный комплекс отбирает у него два электрона и переносит их на убихинон. Убихинон растворим в липидах. Убихинон внутри мембраны диффундирует к комплексу III. Вместе с этим, комплекс I перекачивает 2 протона и 2 электрона из матрикса в межмембранное пространство митохондрий. Перенос электронов от НАДН к О2 включает ряд переносчиков, которые локализованы во внутренней мембране митохондрий. Роль простетической группы играет FMN. С НАДН 2 электрона и протон переносятся на FMN с образованием FMNН2. С FMNH2 электроны переносятся на FeS белки (вторая простетическая группа в молекуле НАДН-дегидрогеназы), от железо-серных центров электроны переносятся на убихинон (кофермент Q). Фермент – НАДН-дегидрогеназа (Оксидоредуктаза, анаэробные дегидрогеназы) Коферменты – FMN, FeS-центры Кофакторы – В2 (рибофлавин) Акцептор электронов – Кофермент Q Энергетический выход – 3 АТФ (т.к передаёт водород в цепь переноса электронов через митохондриальный НАД+) 7. Охарактеризуйте и напишите формулами окислительную реакцию гликолиза: 1) назовите субстраты, продукты, фермент / класс фермента и его группу, кофермент-витамин; 2) охарактеризуйте дальнейшие превращения восстановленного кофермента и объясните механизм работы глицерофосфатного челнока: назовите субстрат для передачи водорода от восстановленного кофермента, фермент и продукты реакции в цитоплазме и митохондриях; 3) объясните последующее окисление восстановленного кофермента в матриксе митохондрий, приводящее к синтезу АТФ: назовите ферменты, коферменты и кофакторы цепи переноса электронов от восстановленного кофермента на конечный акцептор, фермент реакции синтеза АТФ, энергетический выход. 1) Субстрат – Глицеральдегид-3-фосфат Продукт – 1,3 – Бисфосфоглицерат Фермент – Глицеральдегидфосфатдегидрогеназа (класс Оксидоредуктазы, подкласс – Анаэробные дегидрогеназы) Кофермент – НАД+ Витамин – РР/B6 (Никотиновая кислота) 2) НАДН, образующийся при окислении глицеральдегид-3-фосфата в аэробном гликолизе, подвергается окислению путём переноса атомов водорода в митохондриальную дыхательную цепь. Т.к. цитозольный НАДН не может проникнуть в митохондрию, то перенос водорода происходит с помощью челночных систем. 1. Водород от НАДН в цитозоле передается на дигидроксиацетонфосфат ферментом глицерол-3-фосфатдегидрогеназой (НАД-зависимый фермент/флавиновый фермент) с образованием глицерол-3-фосфата. 2. Глицерол-3-фосфат окисляется глицерол-3-фосфатдегидрогеназой (фермент внутренней мембраны митохондрий, ФАД зависимый) в дигидроксиацетонфосфат с образованием ФАДН2 3. Протоны и электроны с ФАДН2 переходят на убихинон и далее по цепи переноса электронов * глицеролфосфатная челночная система работает в клетках белых мышц и гепатоцитов. Однако в клетках сердечных мышц митохондриальная глицерол-3-фосфатдегидрогеназа отсутствует Субстрат для передачи – глицерол-3-фосфат |