Обмен углеводов
 Скачать 1.99 Mb. Скачать 1.99 Mb.
|

|
Ферменты – глицерол-3-фосфатдегидрогеназа (Оксидоредуктазы, анаэробные дегидрогеназы) Продукт: митохондриальный ФАДН2 3) ФАДН2 поступает во 2 комплекс цепи переноса электронов (ФАД-зависимые дегидрогеназы). Он включает в себя ФАД-зависимые ферменты, расположенные на внутренней мембране. Обеспечивает передачу электронов от ФАДН2 на железосерные белки внутренней мембраны митохондрий. Далее эти электроны попадают на коэнзим Q (убихинон). Выход 2 АТФ. Ферменты - ацил-SКоА-дегидрогеназа (β-окисление жирных кислот), сукцинатдегидрогеназа (цикл трикарбоновых кислот), митохондриальная глицерол-3-фосфат-дегидрогеназа (челночный механизм переноса атомов водорода). Класс- оксидоредуктазы, анаэробные дегидрогеназы Кофактор – В2 (рибофлавин) Кофермент - ФАД Энергетический выход – 2 АТФ (т.к водород вводится в цепь переноса электронов на уровне KoQ) 8. Охарактеризуйте процесс полного окисления глюкозы по плану: 1) этапы процесса и их результаты; 2) энергетический выход (объясните полученный результат); 3) аллостерическая регуляция скорости процесса (назовите аллостерические ферменты различных этапов процесса и основные аллостерические активаторы и ингибиторы, объясните механизм аллостерической регуляции); 4) гормональная регуляция скорости процесса (назовите гормон-активатор, ключевые ферменты и возможные способы активации ключевых ферментов при участии данного гормона). 1) Аэробный гликолиз (окисление глюкозы до углекислого газа и воды). Включает в себя несколько стадий: 1. Аэробный гликолиз включает 2 этапа: 1)Подготовительный этап. (1-5 реакции) Глюкоза фосфорилируется и расщепляется на две молекулы фосфотриоз. (Затрачивается 2 молекулы АТФ). 2)Этап, сопряженный с синтезом АТФ. В результате этой серии реакций фосфотриозы превращаются в пируват. Энергия, высвобождающаяся на этом этапе, используется для синтеза 10АТФ. (6-10 реакции) Полученные НАДН поступают в челночные системы. Если в малатаспартатную, то синтезируется 3 АТФ, если в глицерофосфатную, то 2 АТФ. 2. Общий путь катаболизма, включающий превращение пирувата в ацетил-КоА и его окисление в цикле Кребса. В процессе окислительного декарбоксилирования пирувата с участием 3 ферментов (пируватдегидрогеназа, дигидролипоилацетилтрансфераза, дигидролипоилдегидрогеназа) и 5 коферментов (ТПФ, амид липоевой кислоты, Коэнзим А, ФАД и НАД) получается ацетил-КоА. Пируват + НАД+ + HS-KoA -> Ацетил-КоА + НАДН + Н+ + СО2 Ацетил-КоА поступает в цикл Кребса и полностью окисляется до СО2 и Н2О 3. Цепь переноса электронов, сопряженная с реакциями дегидрирования, происходящими в процессе распада глюкозы. 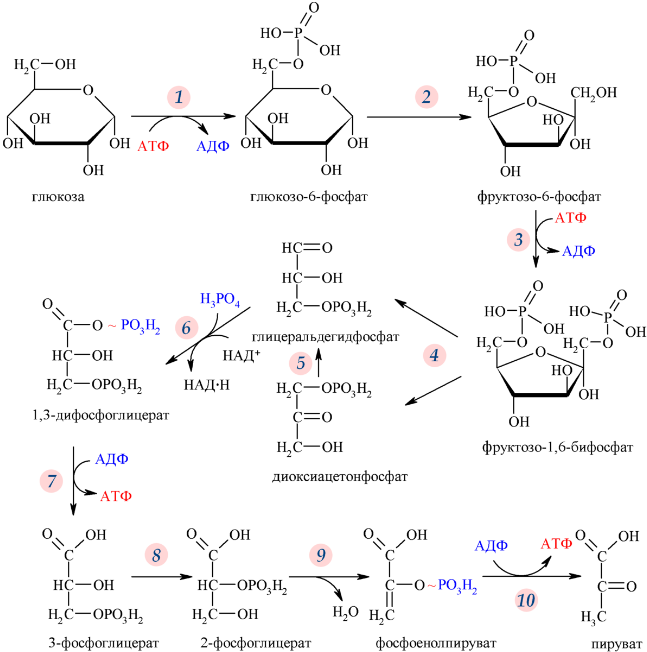 2) 1. Аэробный гликолиз: -2 АТф(подготовительный этап) + 8-10 АТФ (образование пирувата)= 6-8 АТФ 2. Окислительное декарбоксилирование: 2 пирувата -> 2 Ацетил-КоА +6АТФ 3. Цикл Кребса: +24 АТФ Итого 36-38 АТФ. Количество АТФ зависит от того, какой челночный механизм будет включен. Если малат-аспартатный, то из НАДН2 образуется 3 АТФ (3*2=6), если глицерофосфатный, то 2 АТФ (2*2=4). 3) Аллостерические ферменты: Фосфофруктокиназа (3 реакция) - ингибируется АТФ и стимулируется АДФ и АМФ. АТФ в данном случае является и субстратом и аллостерическим регулятором. Трансфераза, фосфотрансфераза Гексокиназа – это аллостерический фермент и ингибируется глюкозой-6-фосфат; она также может фосфорилировать фруктозу, маннозу. Трансфераза, фосфотрансфераза *аллостерическая регуляция - один из типов регуляции активности фермента. Этот тип регуляции представляет собой эффект, наблюдаемый в тех случаях, когда небольшие молекулы (эффекторы), связываясь с ферментом не в области активного центра, изменяют скорость реакции. Подобная регуляция может быть гомотропной, когда молекула субстрата, взаимодействуя с ферментом, изменяет его сродство к молекулам того же субстрата, и гетеротропной, когда сродство к субстрату изменяется при взаимодействии фермента с молекулой, не похожей на молекулы субстрата. Гомотропные и гетеротропные эффекторы могут быть активаторами или ингибиторами. В целом роль аллостерических эффекторов заключается в том, чтобы либо расширить (в случае ингибитора), либо сузить (в случае активатора) диапазон концентраций субстрата, в котором фермент способен увеличивать скорость реакции. 4) 1. Гексокиназа — это регуляторный фермент гликолиза во внепеченочных клетках. Гексокиназа аллостерически ингибируется глюкозо-6-фосфатом. Глюкокиназа — регуляторный фермент гликолиза в гепатоцитах. Синтез глюкокиназы индуцируется инсулином. 2. Фосфофруктокиназа-1. Это главный ключевой фермент, катализирует реакцию, лимитирующую скорость всего процесса (наиболее медленная реакция). Синтез фермента индуцируется инсулином. Аллостерические активаторы — АМФ, АДФ, фруктозо-2,6- дифосфат. Уровень фруктозо-2,6-дифосфата увеличивается под действием инсулина и понижается под действием глюкагона. Аллостерические ингибиторы — АТФ, цитрат. 3. Пируваткиназа - фермент активен в нефосфорилированной форме. Глюкагон (в гепатоцитах) и адреналин (в миоцитах) стимулируют фосфорилирование фермента, а значит инактивируют фермент. Инсулин, наоборот, стимулирует дефосфорилирование фермента, а значит активирует фермент. Аллостерический активатор — Фр-1,6-ФФ. Аллостерический ингибитор — АТФ, ацетил КоА. Синтез фермента индуцирует инсулин. Инсулин. Печень является основным эффекторным органом панкреатических гормонов. Инсулин оказывает множественное действие на печень: он активирует гликолиз, синтез гликогена, жирных кислот и белка, ингибирует глюконеогенез Глюкагон оказывает противоположное инсулину действие. Он стимулирует захват печенью аминокислот, активирует глюконеогенез, ингибирует гликолиз и биосинтез жирных кислот. Глюкагон ингибирует транскрипцию трех регулируемых ферментов гликолиза и активирует продукцию двух ферментов глюконеогенеза. Адреналин ингибирует гликолиз в печени, подобно глюкагону. Однако он оказывает слабовыраженное активирующее действие на гликолиз в мышцах. Кроме этого, адреналин активирует распад жиров (липолиз), гликогена и глюконеогенез. Кортизол ингибирует гликолиз и активирует катаболические процессы в организме: распад гликогена, жиров, белков. 9. Охарактеризуйте процесс регуляции гликолиза и глюконеогенеза в печени в зависимости от ритма питания: 1) назовите ключевые ферменты обоих процессов; 2) назовите гормоны-регуляторы и способы изменения активности ключевых ферментов при участии этих гормонов, а также объясните механизм действия гормонов, приводящий к активации ключевых ферментов; 3) назовите аллостерические ферменты обоих процессов и их аллостерические регуляторы – активаторы и ингибиторы; 4) охарактеризуйте роль бифункционального фермента (БИФ) и фруктозо-2,6-бисфосфата. 1) Ключевые ферменты гликолиза (т.е. те, которые участвуют в необратимых реакциях): 1. Гексокиназа (1 реакция) — это регуляторный фермент гликолиза во внепеченочных клетках. Гексокиназа аллостерически ингибируется глюкозо-6-фосфатом. Глюкокиназа — регуляторный фермент гликолиза в гепатоцитах. Синтез глюкокиназы индуцируется инсулином. 2. Фосфофруктокиназа (3 реакция) -1. Это главный ключевой фермент, катализирует реакцию, лимитирующую скорость всего процесса (наиболее медленная реакция). Синтез фермента индуцируется инсулином. Аллостерические активаторы — АМФ, АДФ, фруктозо-2,6- дифосфат. Уровень фруктозо-2,6-дифосфата увеличивается под действием инсулина и понижается под действием глюкагона. Аллостерические ингибиторы — АТФ, цитрат. 3. Пируваткиназа (10 реакция). Фермент активен в нефосфорилированной форме. Глюкагон (в гепатоцитах) и адреналин (в миоцитах) стимулируют фосфорилирование фермента, а значит инактивируют фермент. Инсулин, наоборот, стимулирует дефосфорилирование фермента, а значит активирует фермент. Аллостерический активатор — Фр-1,6-ФФ. Аллостерический ингибитор — АТФ, ацетил КоА. Синтез фермента индуцирует инсулин. Ключевые ферменты глюконеогенеза: 1. Пируваткарбоксилаза (ПВК: СО2-синтетаза (АТФ→АДФ+Фн)) содержит биотин, находиться в митохондриях, превращает ПВК в ЩУК. Индуктор: глюкагон, адреналин, кортизол. Репрессор: инсулин. Ингибитор: АМФ, активатор АцетилКоА. Образующийся ЩУК проходит внутреннюю мембрану митохондрий в своей восстановленной (в виде малата) или аминоформе (в виде аспартата). 2. Фосфоенолпируваткарбоксикиназа (ГТФ: ЩУК-2-фосфотрансфераза (декарбоксили-рующая)) находиться в цитоплазме, превращает ЩУК в ФЕП. Индуктор: глюкагон, адреналин, кортизол. Репрессор: инсулин. 3. Фруктозо-1,6-фосфотаза (Фруктозо-1,6дф: фосфо-гидролаза) дефосфорилирует фруктозо-1,6дф. Индуктор: глюкагон, адреналин, кортизол. Репрессор: инсулин. Ингибирует АМФ, фруктозо-2,6дф. Активатор: цитрат, жирные кислоты. 4. Глюкозо-6-фосфотаза (Глюкозо-6ф: фосфо-гидролаза) дефосфорилирует глюкозо-6ф. Индуктор: глюкагон, адреналин, кортизол. Репрессор: инсулин. 2) Глюконеогенез: Инсулин индуцирует синтез ключевых ферментов гликолиза и репрессирует синтез ключевых ферментов глюконеогенеза. Глюкагон, кортизол и адреналин индуцирует синтез ключевых ферментов глюконеогенеза. Ключевые ферменты гликолиза активируют – АМФ, фруктозо-2,6дф, фруктозо-1,6дф, ингибируют – АТФ, НАДН2, цитрат, жирные кислоты, аланин, АцетилКоА, глюкагон, адреналин. Ключевые ферменты глюконеогенеза активируют – АцетилКоА, глюкагон, ингибируют – АМФ, фруктозо-2,6дф. Гликолиз: Инсулин. Печень является основным эффекторным органом панкреатических гормонов. Инсулин оказывает множественное действие на печень: он активирует гликолиз, синтез гликогена, жирных кислот и белка, ингибирует глюконеогенез Глюкагон оказывает противоположное инсулину действие. Он стимулирует захват печенью аминокислот, активирует глюконеогенез, ингибирует гликолиз и биосинтез жирных кислот. Глюкагон ингибирует транскрипцию трех регулируемых ферментов гликолиза и активирует продукцию двух ферментов глюконеогенеза. Адреналин ингибирует гликолиз в печени, подобно глюкагону. Однако он оказывает слабовыраженное активирующее действие на гликолиз в мышцах. Кроме этого, адреналин активирует распад жиров (липолиз), гликогена и глюконеогенез. Кортизол ингибирует гликолиз и активирует катаболические процессы в организме: распад гликогена, жиров, белков. 3) Гликолиз: фосфофруктокиназа (3 реакция) – трансферазы, фосфотрансфераза. Она ингибируется АТФ и стимулируется АДФ и АМФ. АТФ в данном случае является и субстратом и аллостерическим регулятором. Гексокиназа – это аллостерический фермент и ингибируется глюкозой – 6 – фосфат; она также может фосфорилировать фруктозу, маннозу. - трансферазы, фосфотрансфераза Глюконеогенез: Пируваткарбоксилаза – аллостерический митохондриальный фермент. Аллостерический активатор – ацетил-КоА. Класс Лигазы Фруктозо-1,6-бисфосфатаза – фермент, ингибируемый АМФ. Класс гидролазы, фосфатазы. 4) Фруктозо-2,6-бисфосфат активирует фосфофруктокиназу и ингибирует фруктозо-1,6-бифосфатазу. Повышение в клетке уровня фруктозо-2,6-бисфосфата способствует усилению гликолиза и уменьшению скорости глюконеогенеза. При снижении концентрации фруктозо-2,6-бисфосфата отмечается обратная картина. Биосинтез и распад фруктозо-2,6- бисфосфата катализируется одним и тем же ферментом, т.е. данный фермент бифункционален, он обладает и фосфокиназной и фосфатазной активностями: Показано также, что бифункциональный фермент в свою очередь регулируется путем цАМФ-зависимого фосфорилирования. Фосфорилирование приводит к увеличению фосфатазной активности и снижению фосфокиназной активности бифункционального фермента. Этот механизм объясняет быстрое действие гормонов, в частности глюкагона, на уровень фруктозо-2,6-бисфосфата в клетке. Активность бифункционального фермента регулируется также и некоторыми метаболитами, среди которых наибольшее значение имеет глицерол-3-фосфат. Действие глицерол-3-фосфата на фермент по своей направленности аналогично эффекту, который наблюдается при его фосфорилировании с помощью цАМФ-зависимых протеинкиназ. 10. Охарактеризуйте пентозофосфатный путь окисления глюкозы по плану: 1) в каких тканях, когда и с какой целью процесс идет наиболее активно; 2) ход реакций: назовите основные этапы и напишите формулами ключевую реакцию (назовите субстрат, фермент / класс фермента и его группу, кофермент-витамин, продукты); 3) результат процесса и его значение; 4) объясните механизм повышения активности ключевого фермента при участии гормона, если известно, что он связан с регуляцией экспрессии гена; назовите гормон-активатор. 1) У млекопитающих активность пентозофосфатного цикла относительно высока в печени, надпочечниках, жировой ткани, эмбриональной ткани и молочной желёз в период лактации. Он поставляет восстановленный НАДФН, необходимый для биосинтеза жирных кислот, холестерина. ПФЦ поставляет пентозофосфаты для синтеза нуклеиновых кислот и многих коферментов. 2) 1 этап. Окислительный 1. дегидрирование глюкозо-6-фосфата при участии фермента глюкозо-6-фосфатдегидрогеназы и кофермента НАДФ+ и образование 6-фосфоглюконолактон 2. гидролиз 6-фосфоглюконолактона с помощью фермента 6-фосфоглюконолактоназы с образованием 6-фосфоглюконата (6-фосфоглюконовая кислота) 3. 6-фосфоглюконат дегидрируется и декарбоксилируется. В результате образуется фосфорилированная кетопентоза – D-рибулозо-5-фосфат и ещё 1 молекула НАДФН 2 этап. Неокислительный (реакции не связаны с использованием кислорода и протекают в анаэробных условиях) РЕАКЦИИ ВЗАИМОПРЕВРАЩЕНИЯ САХАРОВ промежуточный продукты реакций: – рибулозо-5-фосфат – рибозо-5-фосфат – седогептулозо-7-фосфат – фруктозо-6-фосфат – ксилулозо-5-фосфат – глицеральдегид-3-фосфат – эритрозо-4-фосфат Ферменты неокислительной стадии ПФЦ • Трансальдолазы • Транскетолазы Кофермент: тиаминдифосфат Витамин: В1-тиамин Ключевая реакция: Первая реакция–дегидрирование глюкозо-6-фосфата глюкозо-6-фосфатдегидрогеназой с образованием δ-лактона 6-фосфоглюконовой кислоты и НАДФН∙Н+ (НАДФ+ – кофермент глюкозо-6-фосфатдегидрогеназы). 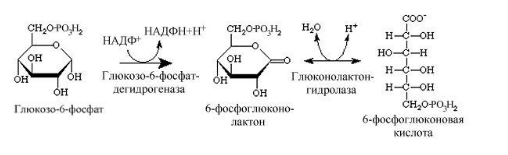 3) Пентозофосфатный цикл обеспечивает организм: 1. Восстановительными эквивалентами (НАДФН). НАДФН обеспечивает: 1. Синтез жирных кислот и других структурных и резервных липидов, 2. Синтез холестерина, желчных кислот, 3. Синтез стероидных гормонов (мужские и женские половые гормоны, кортикостероиды), 4. Синтез витамина D, 5. Микросомальное окисление в печени ядов, лекарственных веществ, эндогенных соединений, 6. Восстановление глютатиона - антиоксиданта 2. Пентозофосфатами (рибозо-5-фосфат) обеспечивает: Синтез нуклеозидов • синтез нуклеотидов (нуклеотидмоно-, ди-, трифосфатов) • синтез нуклеотидных коферментов (НАД, НАДФ, ФАД, КоА) • синтез полимерных производных нуклеотидов - НУКЛЕИНОВЫХ КИСЛОТ (ДНК, РНК, косвенно - синтез БЕЛКА 4) Главными регулируемыми (ключевыми) ферментами пентозофосфатного пути являются две дегидрогеназы его окислительной части: глюкозо-6-фосфатдегидрогеназа и дегидрогеназа 6-фосфоглюконата. Индукторами биосинтеза этих ферментов является инсулин. Активность дегидрогеназ увеличивается при поступлении углеводов в организм и снижается при голодании и диабете. 11. Охарактеризуйте метаболический процесс синтеза гликогена по плану: 1) в каких тканях, когда и с какой целью процесс идет наиболее активно; 2) ход реакций (назовите субстраты, ферменты / класс ферментов, продукты), реакции с затратами АТФ; 3) строение конечного продукта – гликогена; 4) гормональная регуляция скорости процесса (назовите гормон-активатор и объясните механизм его действия, приводящий к синтезу гликогена). 1) Биосинтез гликогена – гликогеногенез происходит в процессе пищеварения. Особенно активно гликогенез происходит в печени и мышцах. Распад гликогена печени служит в основном для поддержания уровня глюкозы в крови в постабсорбтивном периоде. Изменяется в зависимости от ритма питания. Гликоген мышц служит резервом глюкозы - источника энергии при мышечном сокращении. Мышечный гликоген не используется для поддержания уровня глюкозы в крови. В клетках мышц нет фермента глюкозо-6-фосфатазы, и образование свободной глюкозы невозможно. Расход гликогена в мышцах зависит в основном от физической нагрузки . 2) 1-2. Фосфорилирование глюкозы (при участии фермента гексокиназы, а в печени – и глюкокиназы.(ТРАНСФЕРАЗЫ) Далее глюкозо-6-фосфат под влиянием фермента фосфоглюкомутазы переходит в глюкозо-1-фосфат) 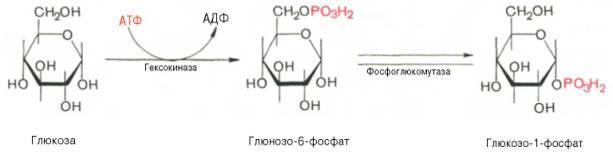 3. Через фосфодиэфирную связь глюкоза соединяется с уридинтрифосфатом, образуя уридиндифосфат-глюкозу и пирофосфат (УДФ–глюкоза является активированной формой глюкозы, непосредственно включающейся в реакцию полимеризации). УДФ – глюкоза переносит глюкозный остаток на затравочное количество гликогена, уже имеющегося в мышцах или печени. Глюкозо-1-фосфат + УТФ < = > УДФ-глюкоза + пирофосфат 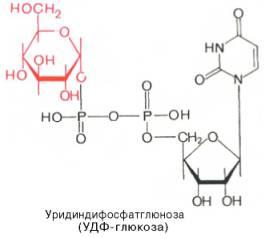 4. Перенос глюкозного остатка, входящего в состав УДФ-глюкозы, на глюкозидную цепь гликогена-праймер. 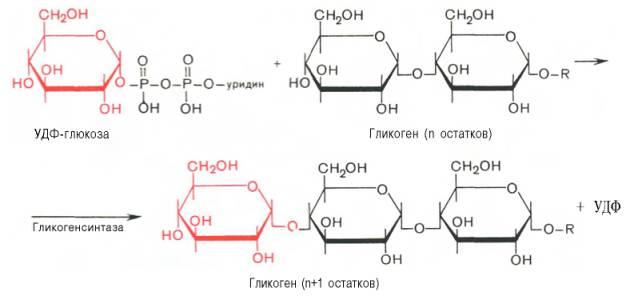 Образующийся УДФ затем вновь фосфорилируется в УТФ за счёт АТФ, и таким образом весь цикл превращений глюкозо-1-фосфата начинается сначала. 3) Гликоген представляет собой разветвленный полисахарид, мономером которого является глюкоза. Остатки глюкозы соединены в линейных участках 1-4 гликозидными связями, а в местах разветвления 1-6 связями 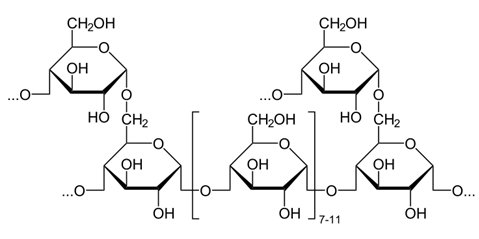 4) Гормон-активатор - инсулин -способствует синтезу гликогена из поступающей в гепатоциты глюкозы путём увеличения транскрипции гена глюкокиназы и активирования гликоген синтазы; 12. Охарактеризуйте метаболический процесс распада гликогена в печени в период голодания по плану: 1) строение гликогена; 2) значение гликогенолиза в печени; 3) ход реакций (назовите субстраты, ферменты / класс ферментов, продукты); 4) гормональная регуляция скорости процесса с характеристикой глюкагона: химическая природа, место синтеза, сигнал для секреции, механизм действия, приводящий к гликогенолизу. 1) Гликоген представляет собой разветвленный полисахарид, мономером которого является глюкоза. Остатки глюкозы соединены в линейных участках 1-4 гликозидными связями, а в местах разветвления 1-6 связями 2) Гликогенолиз — биохимический процесс расщепления гликогена до глюкозы, осуществляется главным образом в печени и мышцах и не требует затрат энергии. Основная задача гликогенолиза — поддержание постоянного уровня глюкозы в крови. Гликоген печени используется главным образом для поддержания физиологической концентрации глюкозы в крови. 3) Распад гликогена происходит путем последовательного отщепления глюкозо-1-фосфата в ответ на повышение потребности организма в глюкозе. Реакцию катализирует гликогенфосфорилаза.(Киназа, фосфорилаза) Образовавшийся в результате фосфоролитического распада гликогена глюкозо-1-фосфат превращается под действием фосфоглюкомутазы в глюкозо-6-фофат. Для осуществления данной реакции необходима фосфорилированная форма фосфоглюкомутазы.  Образование свободной глюкозы из глюкозо-6-фосфата в печени происходит под влиянием глюкозо-6-фосфатазы. Данный фермент катализирует гидролитическое отщепление фосфата: 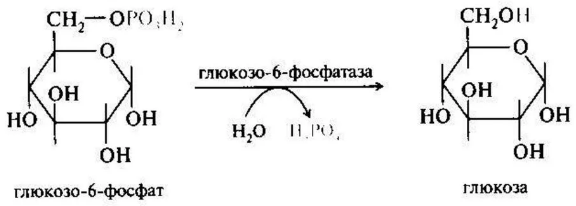 4) Глюкагон образуется из своего предшественника проглюкагона, содержащего на С-конце полипептида дополнительный октапептид (8 остатков), отщепляемый в процессе постсинтетического протеолиза. По биологическому действию глюкагон, как и адреналин, относятся к гипергликемическим факторам, вызывает увеличение концентрации глюкозы в крови главным образом за счет распада гликогена в печени. Органами-мишенями для глюкагона являются печень, миокард, жировая ткань. Биосинтез и секреция глюкагона контролируются главным образом концентрацией глюкозы по принципу обратной связи. Таким же свойством обладают аминокислоты и свободные жирные кислоты. На секрецию глюкагона оказывают влияние также инсулин и инсулиноподобные факторы роста. В механизме действия глюкагона первичным является связывание со специфическими рецепторами мембраны клеток, образовавшийся глюкагонрецепторный комплекс активирует аденилатциклазу и соответственно образование цАМФ. Последний, являясь универсальным эффектором внутриклеточных ферментов, активирует протеинкиназу, которая в свою очередь фосфорилирует киназу фосфорилазы и гликогенсинтазу. Фосфорили-рование первого фермента способствует формированию активной гликоген-фосфорилазы и соответственно распаду гликогена с образованием глюкозо-1-фосфата, в то время как фосфорилирование гликогенсинтазы сопровождается переходом ее в неактивную форму и соответственно блокированием синтеза гликогена. Общим итогом действия глюкагона являются ускорение распада гликогена и торможение его синтеза в печени, что приводит к увеличению концентрации глюкозы в крови.. Установлено, что глюкагон способствует образованию глюкозы из промежуточных продуктов обмена белков и жиров. 13. Охарактеризуйте метаболический процесс распада гликогена в мышцах при физической нагрузке по плану: 1) строение гликогена; 2) значение гликогенолиза в мышцах; 3) ход реакций (назовите субстраты, ферменты / класс ферментов, продукты); 4) гормональная регуляция скорости процесса с характеристикой адреналина: химическая природа, место синтеза, сигнал для секреции, механизм действия, приводящий к гликогенолизу; 5) дополнительные механизмы регуляции гликогенолиза в мышцах при участии кальция и АМФ. 1) Гликоген представляет собой разветвленный полисахарид, мономером которого является глюкоза. Остатки глюкозы соединены в линейных участках 1-4 гликозидными связями, а в местах разветвления 1-6 связями 2) Гликоген мышц служит резервом глюкозы - источника энергии при мышечном сокращении. Мышечный гликоген не используется для поддержания уровня глюкозы в крови. Как уже упоминалось ранее, в клетках мышц нет фермента глюкозо-6-фосфатазы, и образование свободной глюкозы невозможно. Расход гликогена в мышцах зависит в основном от физической нагрузки 3) Распад гликогена происходит путем последовательного отщепления глюкозо-1-фосфата в ответ на повышение потребности организма в глюкозе. Реакцию катализирует гликогенфосфорилаза.(Киназа, фосфорилаза) Образовавшийся в результате фосфоролитического распада гликогена глюкозо-1-фосфат превращается под действием фосфоглюкомутазы в глюкозо-6-фофат. Для осуществления данной реакции необходима фосфорилированная форма фосфоглюкомутазы. *В отличии от печени , мышцы не имеют фермента глюкозы-6-фосфотазы, поэтому глюкозо-6-фосфат используется здесь же, в мышечных клетках, распадаясь аэробным или анаэробным путем. 4) Адреналин вызывает гликогенолиз в печени и липолиз в жировой ткани в присутствии кортизола. Поэтому у адреналэктомированных животных значительно снижены оба эти эффекта адреналина. Вызываемый адреналином гликогенолиз является сложным и многозвеньевым процессом. Он начинается с соединения адреналина с β-адренергическим рецептором на клеточной мембране. Это вызывает активацию аденилатциклазы и приводит к усилению образования циклического аденозинмонофосфата, который, в свою очередь, через ряд реакций приводит к активации фосфорилазы и гликогенолизу. Адреналин влияет также на превращения гликогена в тканях, в частности в мышечной ткани, которая является вторым депо гликогена в организме. Мобилизация гликогена печени и мышц под влиянием адреналина тем интенсивнее, чем богаче ткань организма аскорбиновой кислотой. Если влияние адреналина в печени сводится к глюконеогенетическому действию, то под влиянием адреналина в мышечной ткани усиливается гликогенолиз, в результате которого повышается уровень молочной кислоты. Последняя, выделяясь из ткани в кровь, снова превращается в печени в гликоген. Для того, чтобы ресинтез гликогена мог происходить, часть образовавшейся молочной кислоты должна окисляться. Действительно, введение адреналина сопровождается увеличением газообмена. Это влияние известно как калоригенный эффект адреналина. 5) Существует еще один механизм ускорения мобилизации гликогена при мышечной работе. Киназа фосфорилазы- Са-зависимый фермент.В покоящейся мышце концентрация са в саркоплазме очень низка и киназа фосфорилазы практически неактивна.При поступлении кальция киназа фосфорилазы активируется , кроме того в результате действия аденилаткиназы повышается концентрация АМФ — явлется аллостерическим активатором фосфорилазы б ( обеспечивает скорость мобилизации гликогена , достаточную для выполнения умеренной физической работы) В мышцах фосфорилаза b может активироваться аллостерически под действием АМФ, оставаясь при этом дефосфорилированной. АТФ действует как отрицательный аллостерический эффектор. 14. Охарактеризуйте метаболический процесс синтеза глюкозы (глюконеогенез) по плану: 1) в каких тканях, когда и с какой целью процесс идет наиболее активно; 2) ход реакций синтеза из глицерола (назовите субстраты, ферменты / класс ферментов, коферменты-витамины, продукты); напишите формулами 2 первые реакции, приводящие к образованию дигидроксиацетонфосфата; 3) затраты АТФ; 4) гормональная регуляция скорости процесса при участии глюкагона с характеристикой гормона: химическая природа, место синтеза, сигнал для секреции, механизм действия, приводящий к глюконеогенезу. 1) Глюконеогенез - процесс синтеза глюкозы из веществ неуглеводной природы. Его основной функцией является поддержание уровня глюкозы в крови в период длительного голодания и интенсивных физических нагрузок. Процесс протекает в основном в печени и менее интенсивно в корковом веществе почек, а также в слизистой оболочке кишечника. Механизм глюконеогенеза используется для удаления из крови продуктов тканевого метаболизма, например лактата, образующегося в мышцах и эритроцитах, глицерола, непрерывно образующегося в жировой ткани. 2) Глицерол образуется при гидролизе триацил-глицеролов, главным образом в жировой ткани. Использовать его могут только те ткани, в которых имеется фермент глицерол киназа, например печень, почки. Этот АТФ-зависимый фермент катализирует превращение глицерола в α-глицерофосфат (глицерол-3-фосфат). При включении глицерол-3-фосфата в глюконеогенез происходит его дегидрирование NAD-зависимой дегидрогеназой с образованием дигидроксиацетонфосфата, который далее превращается в глюкозу. 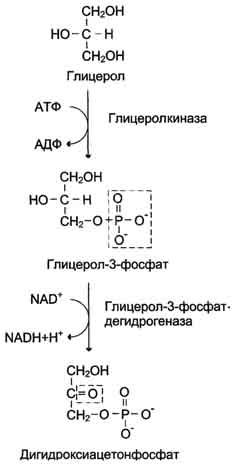 3) 4) Гормональная активация глюконеогенеза осуществляется глюкокортикоидами, которые увеличивают синтез пируваткарбоксилазы, фосфоенолпируват-карбоксикиназы, фруктозо-1,6-дифосфатазы. Глюкагон стимулирует те же самые ферменты через аденилатциклазный механизм путем фосфорилирования. Энергия для глюконеогенеза поступает от β-окисления жирных кислот. Конечный продукт этого окисления, ацетил-КоА аллостерически стимулирует активность первого фермента глюконеогенеза – пируваткарбоксилазы. Кроме этого, фруктозо-1,6-дифосфатаза стимулируется при участии АТФ. 15. Охарактеризуйте метаболический процесс синтеза глюкозы (глюконеогенез) по плану: 1) в каких тканях, когда и с какой целью процесс идет наиболее активно; 2) ход реакций синтеза из аланина (назовите субстраты, ферменты / класс ферментов, коферменты-витамины, продукты); напишите формулами первую реакцию; 3) затраты АТФ; 4) гормональная регуляция скорости процесса при участии кортизола с характеристикой гормона: химическая природа, место синтеза, сигнал для секреции, механизм действия, приводящий к глюконеогенезу. 1) Глюконеогенез - процесс синтеза глюкозы из веществ неуглеводной природы. Его основной функцией является поддержание уровня глюкозы в крови в период длительного голодания и интенсивных физических нагрузок. Процесс протекает в основном в печени и менее интенсивно в корковом веществе почек, а также в слизистой оболочке кишечника. Механизм глюконеогенеза используется для удаления из крови продуктов тканевого метаболизма, например лактата, образующегося в мышцах и эритроцитах, глицерола, непрерывно образующегося в жировой ткани. 2) Аланин, трансаминируясь, образует пировиноградную кислоту, которая способна карбоксилироваться до оксалоацетата. Оксалоацетат, являясь первым элементом в процессе глюконеогенеза, далее включается в состав глюкозы. Из всех аминокислот, поступающих в печень, примерно 30% приходится на долю аланина. Это объясняется тем, что при расщеплении мышечных белков образуются аминокислоты, многие из которых превращаются сразу в пируват или сначала в оксалоацетат, а затем в пируват. Последний превращается в аланин, приобретая аминогруппу от других аминокислот. Аланин из мышц переносится кровью в печень, где снова преобразуется в пируват, который частично окисляется и частично включается в глюкозонеогенез. Следовательно, существует следующая последовательность событий (глюкозо-аланиновый цикл): глюкоза в мышцах → пируват в мышцах → аланин в мышцах → аланин в печени → глюкоза в печени → глюкоза в мышцах . Весь цикл не приводит к увеличению количества глюкозы в мышцах, но он решает проблемы транспорта аминного азота из мышц в печень и предотвращает лактоацидоз. 1. 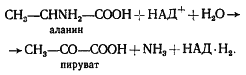 2. 2. 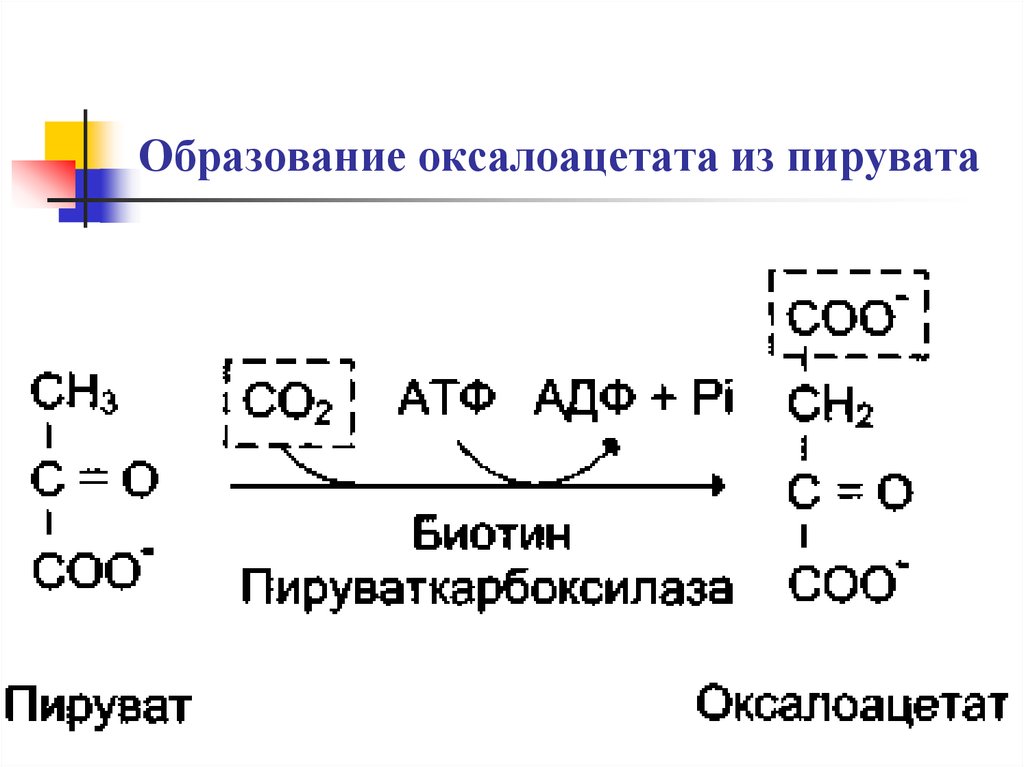 3. 3.  3) В ходе этого процесса расходуется 6 моль АТФ на синтез 1 моль глюкозы из 2 моль пирувата. 4 АТФ расходуется на стадии синтеза фосфоенолпирувата из оксалоацетата и еще 2 моль АТФ на стадиях образования 1,3-бисфосфоглицерата из 3-фосфоглицерата. Суммарный результат глюконеогенеза из пирувата выражается следующим уравнением: 2 Пируват + 4АТФ + 2ГТФ + 2(NАDН+Н+) + 4H2O → Глюкоза + 4АДФ + 2ГДФ + 6Н3РO4+2NАD+. 4) Кортизол — биологически активный глюкокортикоидный гормон стероидной природы, то есть в своей структуре имеет стерановое ядро. Биосинтез. Кортизол синтезируется корой надпочечников из холестерола.У взрослого человека надпочечники за сутки выделяют 15-30 мг кортизола. Скорость синтеза и секреции кортизола стимулируются в ответ на стресс, травму, инфекцию, понижение концентрации глюкозы в крови. Повышение концентрации кортизола подавляет синтез кортиколиберина и АКТГ по механизму отрицательной обратной связи. Нервная система реагирует на внешние воздействия (в том числе стрессовые), посылая нервные импульсы в гипоталамус. В ответ на сигнал гипоталамус секретирует кортиколиберин, который переносится кровью по т.н. воротной системе прямо в гипофиз и стимулирует секрецию им АКТГ. Последний поступает в общий кровоток и, попав в надпочечники, стимулирует выработку и секрецию корой надпочечников кортизола. Выделившийся в кровь кортизол достигает клеток-мишеней (в частности, клеток печени), проникает путем диффузии в их цитоплазму и связывается там со специальными белками – рецепторами кортизола. Образовавшиеся гормон-рецепторные комплексы после «активации» связываются с соответствующей областью ДНК и активируют определенные гены, что в итоге приводит к увеличению выработки специфических белков. Именно эти белки и определяют ответную реакцию организма на кортизол, а значит, и на внешнее воздействие, послужившее причиной его секреции. Реакция состоит, с одной стороны, в усилении синтеза глюкозы в печени и в проявлении (разрешении) действия многих других гормонов на обменные процессы, а с другой – в замедлении распада глюкозы и синтеза белков в ряде тканей, в том числе мышечной. Таким образом, эта реакция направлена в основном на экономию имеющихся энергетических ресурсов организма (снижение их расходования мышечной тканью) и восполнение утраченных: синтезируемая в печени глюкоза может запасаться в виде гликогена – легко мобилизуемого потенциального источника энергии. 16. Охарактеризуйте основной путь метаболизма галактозы в печени, приводящий к образованию гликогена и глюкозы, по плану: 1) поступление галактозы с пищей в составе лактозы и усвоение в ЖКТ; 2) ход реакций, приводящих к образованию гликогена в печени (назовите субстраты, ферменты / класс ферментов, продукты); 3) ход реакций, приводящих к образованию глюкозы в печени (назовите субстраты, ферменты / класс ферментов, продукты). 1) Галактоза образуется в кишечнике в результате гидролиза лактозы. Молочный сахар — лактоза содержится только в молоке. Гидролиз лактозы происходит в кишечнике, причем протекает он медленно, в связи с чем процессы брожения в кишечнике ограничиваются и нормализуется жизнедеятельность полезной кишечной микрофлоры. 2) Подготовительные этапы: 1 этап. Фософорилирование глюкозы ГЛЮКОЗА+АТФ->ГЛЮКОЗО-6-ФОСФАТ+АДФ Фермент:глюкокиназа(гексокиназа) транфераза 2 этап.Фосфоглюкомутазная реакция ГЛЮКОЗО-6-ФОСФАТ->ГЛЮКОЗО-1-ФОСФАТ фермент:фосфоглюкомутаза трансфераза Синтез гликогена: 1-я стадия синтеза: ГЛЮКОЗО-6-ФОСФАТ+УТФ->УДФ-ГЛЮКОЗА+PP фермент:УДФ-глюкопирофосфорилаза транфераза 2-я стадия синтеза: УДФ-ГЛЮКОЗА + (C6H10O5)n ->УДФ+(C6H10O5)n+1 фермент:гликогенситаза трансфераза (C6H10O5)n+1 - это и есть гликонен Так как гликоген в клетке никогда не расщепляется полностью, синтез гликогена осуществляется путём удлинения уже имеющейся молекулы полисахарида, называемой "затравка", или "праймер". К "затравке" последовательно присоединяются молекулы глюкозы. Строением молекулы "затравки" как бы предопределяется тип связи, который возникает в реакции трансгли-козилирования. Таким образом, синтезируется полисахарид, аналогичный по строению с "затравочным" Ветвящий фермент: амило-1,4->1,6-трансглюкозидаза -переносит фрагмент цепи (6 остатков глюкозы) из линейной цепи и присоединяет его a-1,6-глюкозидной связью. 3) Галактоза образуется в кишечнике в результате гидролиза лактозы. Чтобы превратить галактозу в глюкозу, необходимо изменить оптическую конфигурацию Н- и ОН-групп С4 атома в галактозе(реакция эпимеризации). Эта реакция возможна только с УДФ-производным галактозы. УДФ-галактоза образуется из УДФ-глюкозы. Включение галактозы в описанную реакцию. Эпимеризации предшествует ее фосфорилирование с образованием галактозо-1-фосфата. Далее галактозо-1-фосфат замещает остаток глюкозы в УДФ-глюкозе с образованием УДФ-галактозы(2 реакция). Затем галактоза в составе нуклеотида включается в реакцию эпимеризации, в которой участвует эпимераза – NAD-зависимый фермент, катализирующий окисление и восстановление галактозы по С4 углеродному атому Метаболизм галактозы в норме: 1)галактоза + АТФ = галактозо-1-фосфат + АДФ (фермент –галактокиназа : класс – трансфераза; субстрат - галактоза) 2)галактозо-1-фосфат + УДФ-глюкоза = глюкозо-1-фосфат + УДФ-галактоза (фермент –гексозо-1-фосфат-уридилтрансфераза : класс- трансферазы) 3)УДФ-галактоза = УДФ-глюкоза (фермент: галактозо-УДФ-эпимераза : класс- изомеразы). 17. Лечение больных с галактоземией заключается в исключении из питания продуктов, содержащих галактозу. Однако галактоза необходима в организме, например, для синтеза протеогликанов – компонентов соединительной ткани. Образование галактозы в организме возможно из глюкозы. Охарактеризуйте: 1) причины галактоземии; 2) последовательность реакций (назовите субстраты, ферменты / классы ферментов, продукты), приводящих к образованию галактозы из глюкозы, учитывая, что субстратом изомеразы является УДФ-глюкоза; напишите формулы глюкозы и галактозы. 1) Галактоземия - наследственная ферментопатия, характеризующаяся нарушением нормального процесса углеводного обмена, а именно – метаболизма галактозы. Признаками галактоземии являются непереносимость грудного молока и молочных смесей, рвота, анорексия, гипотрофия, желтуха, цирроз печени, спленомегалия, отеки, катаракта, задержка психомоторного развития. Галактоземия возникает при нарушении обмена галактозы, обусловленном наследственным дефектом любого из 3 ферментов, включающих галактозу в метаболизм глюкозы (Галактокиназа, Галактозо-1-фосфат-уридилтрансфераза, уридилфосфат-4-эпимераза). 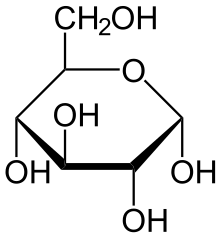 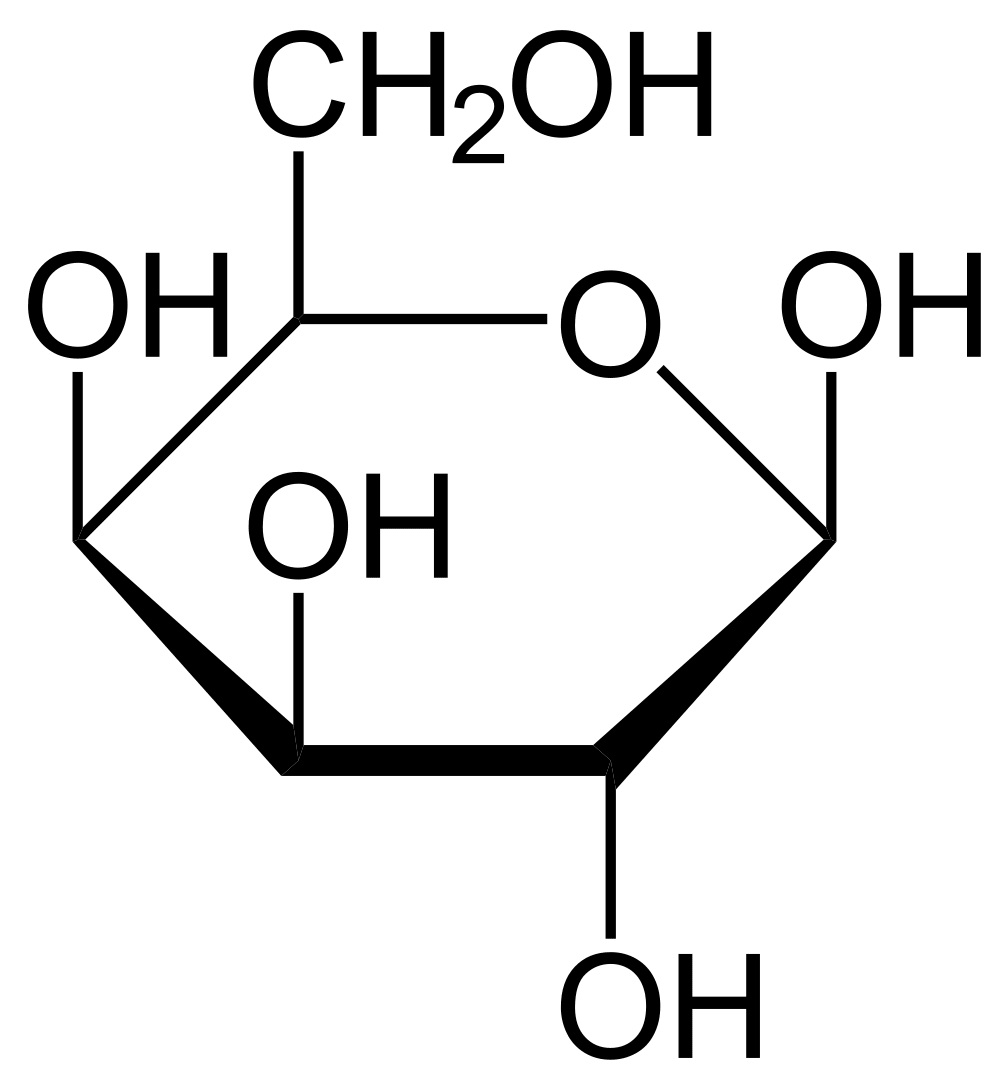 Глюкоза Галактоза 2) 1)глюкоза + АТФ = глюкозо-6-фосфат + АДФ (фермент – гексониказа : класс – трансфераза) 2)глюкозо-6-фосфат = глюкозо-1-фосфат ( фермент – фосфоглютомутаза : класс-внутриклеточн. Трансферазы) 3)УДФ-глюкоза = УДФ-галактоза (глюкозо-УДФ-эпимераза : класс-изомеразы) 4) глюкозо-1-фосфат + УДФ-галактоза = галактозо-1-фосфат + УДФ-глюкоза (ферметн – глюкозо-1-фосфат-уридилтрансфераза) 5)галактазо-1-фосфат + АДФ = галактоза + АТФ 18. Охарактеризуйте основной путь метаболизма фруктозы в печени с участием фруктокиназы, приводящий к образованию глюкозы, по плану: 1) поступление с пищей в составе сахарозы и усвоение в ЖКТ; 2) ход реакций, приводящих к образованию глюкозы в печени (назовите субстраты, ферменты / класс ферментов, продукты); напишите формулами реакцию фруктокиназы. Почему больным сахарным диабетом рекомендуют заменить глюкозу на фруктозу? 1) В отличие от глюкозы и сахарозы фруктоза не требует в организме человека воздействия инсулина – гормона, который обязателен при расщеплении глюкозы или обычного сахара. Фруктоза усваивается напрямую некоторыми клетками нашего организма - мужскими половыми клетками – сперматозоидами и клетками печени Фруктоза, напрямую расщепляясь клетками печени, дальше превращается в жирные кислоты, которые затем организмом превращаются в жир. Значительное количество фруктозы, образующееся при расщеплении сахарозы, прежде чем поступить в систему воротной вены, превращается в глюкозу уже в клетках кишечника. Другая часть фруктозы всасывается с помощью белка-переносчика, т.е. путём облегчённой диффузии. 2) Метаболизм фруктозы начинается с реакции фосфорилирования (реакция 1), катализируемой фруктокиназой с образованием фруктозо-1-фосфата. Фермент обнаружен в печени, а также в почках и кишечнике. Этот фермент обладает абсолютной специфичностью, поэтому, в отличие от глюкокиназы, инсулин не влияет на его активность. Последнее обстоятельство объясняет, почему уровень выведения фруктозы в моче у больных сахарным диабетом и здоровых не отличается. Фруктоза-1-фосфат не может превращаться во фруктозо-6-фосфат из-за отсутствия соответствующего фермента. Вместо этого фруктозо-1-фосфат далее расщепляется фруктозо-1-фосфатальдолазой (альдолаза В) на глицеральдегид и дигидроксиацетон-3-фосфат (реакция 2). Последний является промежуточным продуктом гликолиза и образуется в ходе реакции, катализируемой фруктозо-1,6-бисфосфосфатальдолазой (альдолаза А). Глицеральдегид может включаться в гликолиз после его фосфорилирования с участием АТФ (реакция 3). Две молекулы триозофосфатов либо распадаются по гликолитическому пути, либо конденсируются с образованием фруктозо-1,6-бисфосфата и далее участвуют в глюконеогенезе (реакции 8, 7, 5, 9). Фруктоза в печени включается главным образом во второй путь. Часть дигидроксиацетон-3-фосфата может восстанавливаться до глицерол-3-фосфата и участвовать в синтезе триацилглицеролов. Следует отметить, что включение фруктозы в метаболизм через фруктозо-1-фосфат минует стадию, катализируемую фосфофруктокиназой (реакция 6), которая является пунктом метаболитического контроля скорости катаболизма глюкозы. Этим обстоятельством можно объяснить, почему увеличение количества фруктозы ускоряет в печени процессы, ведущие к синтезу жирных кислот, а также их этерификацию с образованием триацилглицеролов. 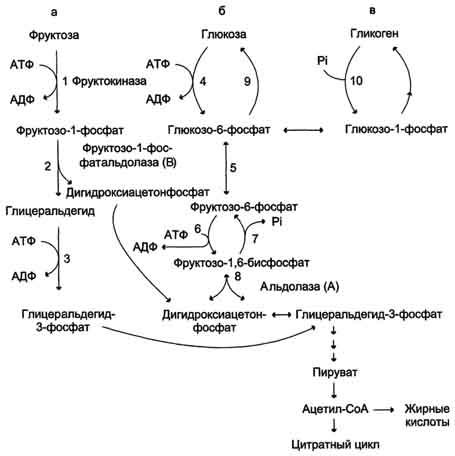 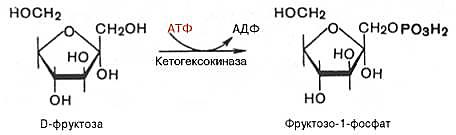 *В отличие от глюкозы и сахарозы фруктоза не требует в организме человека воздействия инсулина – гормона, который обязателен при расщеплении глюкозы или обычного сахара. Фруктоза, напрямую расщепляясь клетками печени, дальше превращается в жирные кислоты, которые затем организмом превращаются в жир. А из-за того, что фруктоза не повышает уровень глюкозы в крови, человек сильнее и чаще испытывает чувство голода и начинает больше есть. 19. Охарактеризуйте дополнительный путь метаболизма фруктозы с участием фруктокиназы, протекающий по гликолитическому пути, по плану: 1) поступление с пищей в составе сахарозы и усвоение в ЖКТ; 2) ход реакций гликолитического расщепления, приводящих к образованию пирувата или лактата (назовите субстраты, ферменты / классы ферментов, продукты); напишите формулами реакцию фруктокиназы. Почему расщепление фруктозы по гликолитическому пути происходит быстрее, чем расщепление глюкозы? 1) Значительное количество фруктозы, образующееся при расщеплении сахарозы, прежде чем поступить в систему воротной вены, превращается в глюкозу уже в клетках кишечника. Другая часть фруктозы всасывается с помощью белка-переносчика, т.е. путём облегчённой диффузии. 2) *Метаболизм фруктозы начинается с реакции фосфорилирования (реакция 1), катализируемой фруктокиназой с образованием фруктозо-1-фосфата. Фруктокиназа/кетогексокиназа - Фосфотрансферазы/Киназы - Фермент обнаружен в печени, а также в почках и кишечнике; обладает абсолютной специфичностью, поэтому, в отличие от глюкокиназы, инсулин не влияет на его активность. (поэтому уровень выведения фруктозы в моче у больных сахарным диабетом и здоровых отличается) *Фруктозо-1-фосфат не может превращаться в фруктозо-6-фосфат из-за отсутствия соответствующего фермента. Вместо этого фруктозо-1-фосфат далее расщепляется фруктозо-1-фосфатальдолазой (альдолаза В) на глицеральдегид и дигидроксиацетон-3-фосфат (реакция 2). Дигидроксиацетон-3-фосфат является промежуточным продуктом гликолиза и образуется в ходе реакции, катализируемой фруктозо-1,6-бисфосфосфатальдолазой (альдолаза А). Фруктозо-1-фосфатальдолаза - Лиазы *Глицеральдегид может включаться в гликолиз после его фосфорилирования с участием АТФ (реакция 3). *Две молекулы триозофосфатов либо распадаются по гликолитическому пути, либо конденсируются с образованием фруктозо-1,6-бисфосфата и далее участвуют в глюконеогенезе (реакции 8, 7, 5, 9). Фруктоза в печени включается главным образом во второй путь. *Часть дигидроксиацетон-3-фосфата может восстанавливаться до глицерол-3-фосфата и участвовать в синтезе триацилглицеролов. *Следует отметить, что включение фруктозы в метаболизм через фруктозо-1-фосфат минует стадию, катализируемую фосфофруктокиназой (реакция 6), которая является пунктом метаболитического контроля скорости метаболизма глюкозы. Этим обстоятельством можно объяснить, почему увеличение количества фруктозы ускоряет в печени процессы, ведущие к синтезу жирных кислот, а также их этерификацию с образованием триацилглицеролов. Фосфофруктокиназа - Фосфотрансферазы/Киназы 20. Охарактеризуйте последовательность реакций (назовите субстраты, ферменты / классы ферментов, продукты) образования гликогена в печени из исходного субстрата фруктозы в 2-х вариантах: 1) с участием гексокиназы, которая катализирует реакцию фосфорилирования по 6 атому углерода; 2) с участием фруктокиназы, которая катализирует реакцию фосфорилирования по 1 атому углерода, и альдолазы фруктозо-1-фосфата. В целом переход фруктозы в глюкозу осуществляется по двум направлениям. Сначала происходит активация фруктозы посредством фосфорилирования либо 6-го атома углерода при участии гексокиназы, либо 1-го атома при участии фруктокиназы. В печени имеются оба фермента, однако гексокиназа имеет гораздо более низкое сродство к фруктозе и в ней этот путь слабо выражен. Образованный ею фруктозо-6-фосфат далее изомеризуется и глюкозо-6-фосфатаза отщепляет уже ненужный фосфат с получением глюкозы. 1) В печени имеются оба фермента, однако гексокиназа имеет гораздо более низкое сродство к фруктозе и в ней этот путь слабо выражен. Образованный ею фруктозо-6-фосфат далее изомеризуется и глюкозо-6-фосфатаза отщепляет уже ненужный фосфат с получением глюкозы. 1-2. Фосфорилирование глюкозы (при участии фермента гексокиназы, а в печени – и глюкокиназы.(ТРАНСФЕРАЗЫ) Далее глюкозо-6-фосфат под влиянием фермента фосфоглюкомутазы переходит в глюкозо-1-фосфат) 3. Через фосфодиэфирную связь глюкоза соединяется с уридинтрифосфатом, образуя уридиндифосфат-глюкозу и пирофосфат (УДФ–глюкоза является активированной формой глюкозы, непосредственно включающейся в реакцию полимеризации). УДФ – глюкоза переносит глюкозный остаток на затравочное количество гликогена, уже имеющегося в мышцах или печени. |